Nedsatt epitel ion transport til grunn organisme ione- og osmoregulering. Drosophila melanogaster Malpighian (nedsatt) tubuli tilbyr en enestående mulighet til å studere de molekylære mekanismene for epitel ion transport. Dette skyldes kombinasjonen av kraftige genetikk av Drosophila, sammen med tilgjengeligheten av sine nyretubuli til fysiologisk undersøkelse. The Ramsay analysen, oppkalt etter etterforsker som pioner teknikken en måler fluidsekresjon priser fra isolerte Malpighian tubuli, og ble etablert i Drosophila i 1994 av Dow og kolleger 2. Dette banet vei for videre studier ved hjelp Drosophila genetiske verktøy, for eksempel GAL4-UAS system 3,4, for å definere celle-spesifikke signalveier som regulerer fluidsekresjon. Et eksempel inkluderer kalsiumsignalisering i respons til et peptidhormon 5, blant mange andre 6,7.
ve_content "> En kombinasjon av genetiske teknikker og klassiske fysiologiske studium har vist at urinproduksjonen i fore fly gjennom sekresjon av et kaliumklorid-rik væske fra den største delen av rørelementet. Dette skjer gjennom parallell transepitelial sekresjon av kationer, i første rekke K + Na +, men også gjennom hovedcellen, og Cl -. sekresjon gjennom stel cellen 8-12 Muligheten for separat å måle transepitelial K + og Na + fluks gir en mer detaljert karakterisering av transportmekanismer enn måling av fluidsekresjon alene. For eksempel, i ikke-stimulerte Drosophila tubuli, Na + / K + -ATPase-inhibitoren ouabain har ingen virkning på væske sekresjon 2, selv når dens opptak i primære celler inhiberes av det organiske anion transporter inhibitor taurocholat 13. Imidlertid, og Linton O'Donnell viste at ouabain depolarisererden basolaterale membran potensial, og øker Na + fluks 9. Som vist i de representative resultater, vi kopiert disse funnene, og viste at K + flux er samtidig redusert 14; økt Na + flux og redusert K + flux har motstridende effekter på fluidsekresjon, noe som resulterer i ingen netto endring i sekret. Således er det to oppløsninger til de "ouabain paradoks", dvs. den opprinnelige observasjonen som ouabain har noen effekt på fluidsekresjon i Drosophila tubuli. Første, i stimulerte tubuli, er ikke tydelig effekten av ouabain på fluidsekresjon grunn dens opptak i det organiske anion transporter 13; og andre, i unstimulated tubuli, ouabain har motstridende effekter på transepitelial Na + og K + flux, som resulterer i ingen netto endring i fluidsekresjon (se Representative Resultater og ref. 9). Derfor den primære rolle Na + / K + -ATPase i ustimulerte tubuli er å senke intracellulær Na + -konsentrasjon å generere en gunstig konsentrasjonsgradient for Na + -koblet transportprosesser over basolaterale membran. Faktisk, ved separat måling Na + og K + flukser, viste vi at tubuli mangler fly natrium-kalium-to-klorid kotransporter (NKCC) har sunket transepitelial K + flux, uten ytterligere nedgang etter ouabain tillegg og ingen endring i transepitelial Na + flux 14. Disse funnene støtter vårt konklusjon at Na + inn i cellen gjennom NKCC resirkuleres via Na + / K + -ATPase. I et annet eksempel Ianowski et al., Observert at senking bad K + konsentrasjon fra 10 mM til 6 mM redusert transepitelial K + fluks og øket transepitelial Na + fluksen i tubuli fra Rhodnius prolixus, med ingen netto endring i fluidsekresjon <sup> 15. Forskjellige effekter på Na + flux og K + flux over larve tubuli har også blitt observert i Drosophila tubuli i respons til varierende salt dietter 16 og i to myggarter som svar på stell saltholdighet 17.Den største utfordringen i målingen av transepitelial ionefluks i Ramsay analysen forberedelse er bestemmelse av ionekonsentrasjoner innenfor utskilt væske. Denne utfordringen har blitt møtt med varierende løsninger, inkludert flamme photometery 18, bruk av radioaktive ioner 19, og elektron probe bølgelengde spredning spektroskopi 20. Disse teknikkene krever overføring av det utskilte fluid fallet til et instrument for måling av ion-konsentrasjoner. Siden volumet av væske som utskilles av den ustimulerte Drosophila tubuli er liten, typisk ~ 0,5 nl / min, utgjør dette en teknisk utfordring og også introduserer feil hvis noe av det utskilte fluid ertapt ved overføring. I motsetning til dette tillater bruken av ione-spesifikke elektroder til måling av ioneaktivitet (som ion-konsentrasjon kan beregnes) in situ. Den nåværende protokoll ble tilpasset fra det som brukes av Maddrell og kolleger til å måle transepitelial K + fluks over Rhodnius tubuli ved hjelp valinomycin som ionofor K + 21, og også beskriver bruk av en 4-tert–butylcalix [4] aren-tetraeddiksyre tetraetyl-ester-baserte Na + -spesifikk ione-spesifikke elektroder kjennetegnet ved Messerli et. al. 22. Ion-spesifikke elektroder har også blitt brukt til å måle ionekonsentrasjoner i væske som skilles ut av Malpighian tubuli i Ramsay analysen i voksen 9,23 og larve 16 Drosophila melanogaster, New Zealand Alpine Weta (Hemideina maori) 24 og mygg 17.
Her beskriver vi i detalj ved bruk av Ramsay såsi å måle fluid sekresjon priser i Malpighian tubuli fra Drosophila melanogaster, samt anvendelse av ione-spesifikke elektroder for å bestemme konsentrasjonene av K + og Na + i den utskilte væske og dermed beregningen av transepitelial ion flukser. En oversikt av analysen er gitt i figur 1.
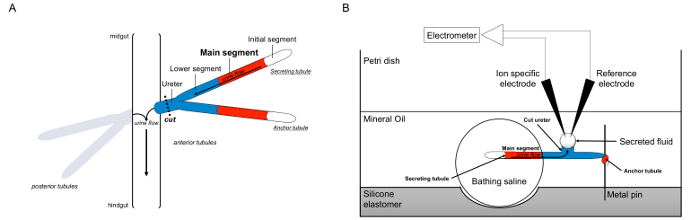
Figur 1. Skjematisk av Malpighian Drikkerør og Ramsay analysen med bruk av Ion-spesifikke Elektroder måle ionekonsentrasjoner. Denne figuren illustrerer oppsettet for Ramsay analysen. (A) Hver flue har fire rør, et par av fremre tubuli og et par bakre rør, som flyter i bukhulen omgitt av hemolymph. I hvert par, de to tubuli delta på ureter, som munner da urinen i krysset midgut og hindgut. Rørelementene er blind-avsluttet. Urin genereres av fluid-sekresjon hovedsegmentet (vist i rødt), og strømmer mot ureter og ut i tarmen. Etter disseksjon, er tubule paret skilt fra tarmen ved transecting ureter. (B) De to rørelementene blir deretter overført til en dråpe av badesaltløsning inne i en brønn av analysen fatet. En av de to rørelementene, referert til her som "anker tubuli," er viklet rundt en metallstift, og er inert. Den andre tubuli er sekresjon tubuli. Den innledende segment (som ikke skiller ut væske) og hovedsegmentet av de som utskiller tubuli forblir i dråpen av badesaltløsning. Ioner og vann flytte fra bade saltvann og inn i tubulære lumen av hovedsegmentet, og deretter bevege seg mot ureter, som ville oppstå in vivo. Den nedre segment (blå) er utenfor bade saltvann og derfor inert. Siden ureter er kuttet, framgår det utskilte fluid som en dråpe fra den kappede enden av ureter. THan utskilt væske dråpe forstørrer over tid som sekresjon fortsetter, og dens diameter er målt ved hjelp av et okular mikrometer. Et lag av mineralolje hindrer fordampning av det utskilte fluid. Referanse- og ion spesifikke elektroder måler ion konsentrasjonen av utskilt væske. Klikk her for å se en større versjon av dette tallet.
