Transporte de íons epitelial renal subjacente iono- organismal e osmoregulation. A Drosophila melanogaster Malpighian (renal) túbulo oferece uma oportunidade única para estudar os mecanismos moleculares de transporte de íons epitelial. Isto é devido à combinação dos poderosas da genética de Drosophila, emparelhado com a acessibilidade dos seus túbulos renais para estudo fisiológico. O ensaio de Ramsay, em homenagem ao pesquisador que foi pioneiro na técnica 1, mede as taxas de secreção de fluidos de túbulos de Malpighi isolados, e foi criada em Drosophila em 1994 pela Dow e colegas 2. Isso pavimentou o caminho para novos estudos utilizando Drosophila ferramentas genéticas, tais como o sistema GAL4-UAS 3,4, para definir específico de células vias de sinalização que regulam a secreção de fluidos. Um exemplo inclui a sinalização de cálcio, em resposta a uma hormona peptídica, 5, 6,7, entre muitos outros.
ve_content "> uma combinação de técnicas genéticas e estudo fisiológico clássica demonstrou que a geração de urina na mosca ocorre através da secreção de um líquido rico em cloreto de potássio a partir do segmento principal do túbulo. Isto ocorre através da secreção transepitelial paralelo de catiões, principalmente K + Na +, mas também, através da célula principal, e Cl -. através da secreção de células estreladas 8-12 A capacidade de medir separadamente transepiteliais K + e Na + fluxos permite uma caracterização mais detalhada dos mecanismos de transporte do que a medição da secreção de fluido sozinho. Por exemplo, nos túbulos de Drosophila não estimuladas, a Na + / K + -ATPase ouabaína não tem efeito sobre a secreção de fluido 2, mesmo quando a sua absorção em células principais é inibida pelo inibidor de transportador aniónico taurocolato orgânica 13. No entanto, e Linton O'Donnell mostrou que a ouabaína despolarizao potencial da membrana basolateral, e aumenta o fluxo de Na + 9. Como mostrado nos resultados representativos, nós replicado estes achados, e mostrou que K + fluxo é concomitantemente diminuiu 14; o aumento do fluxo de Na + e diminuição do K + fluxo têm efeitos opostos sobre a secreção de fluido, resultando em nenhuma alteração na secreção de líquido. Assim, existem duas resoluções para o "paradoxo ouabaína," ou seja, a observação inicial de que a ouabaína não tem efeito sobre a secreção de fluido no túbulo Drosophila:. Em primeiro lugar, nos túbulos estimulada, o efeito de ouabaína na secreção de fluido não é aparente devido à sua absorção pelo transportador de aniões orgânicos 13; e, segundo, nos túbulos não estimuladas, ouabaína tem efeitos sobre transepithelial Na + e K + fluxo opostas, resultando em nenhuma alteração líquida na secreção de fluidos (ver resultados representativos e ref. 9). Portanto, o principal papel do Na + / K + -ATPase nos túbulos não estimuladas é o de diminuir a concentração intracelular de Na + para gerar um gradiente de concentração de Na + para o favorável -coupled processos de transporte através da membrana basolateral. Com efeito, medindo em separado de Na + e K + fluxos, nós demonstramos que túbulos que falta o co-transportador de mosca de sódio-potássio-2-cloreto de (CCAN) diminuíram K transepitelial + fluxo, sem diminuição adicional após a adição de ouabaína, e nenhuma alteração em transepitelial Na + fluxo 14. Estes resultados suportada nossa conclusão de que o Na + que entra na célula através da CCAN é reciclado através do Na + / K + -ATPase. Em outro exemplo, Ianowski et al., Observaram que a redução da concentração do banho de K + de 10 mM a 6 mM de K + diminuiu transepitelial fluxo transepitelial e aumentou o fluxo de Na + nos túbulos de Rhodnius prolixus, sem alteração líquida no fluido de secreção <sup> 15. Efeitos diferenciais de fluxo de Na + e K + fluxo através dos túbulos de larvas também têm sido observadas em túbulos de Drosophila em resposta a diferentes dietas de sal 16 e em duas espécies de mosquitos em resposta à criação de salinidade 17.O maior desafio na medição de fluxo de iões transepitelial na preparação de ensaio Ramsay é a determinação das concentrações de iões dentro do fluido segregado. Este desafio foi cumprido com soluções diferentes, incluindo photometery chama 18, o uso de íons radioativos 19, e eletrônica sonda comprimento de onda espectroscopia por dispersão de 20. Estas técnicas exigem transferência da gota líquido secretado a um instrumento para a medição de concentrações de íons. Uma vez que o volume de fluido que é segregado pelo túbulo Drosophila não estimulado é pequena, tipicamente ~ 0,5 nl / min, isto representa um desafio técnico e também introduz erro se algum do fluido é segregadaperdeu por ocasião da transferência. Em contraste, a utilização de eléctrodos específicos de iões permite a medição da actividade de iões (a partir do qual a concentração de iões pode ser calculado) in situ. O protocolo corrente foi adaptada da utilizada por Maddrell e colegas para medir transepitelial K + fluxo através do túbulo Rhodnius usando valinomicina como o K + ionóforo 21, e também descreve a utilização de um -butylcalix 4- terc [4] areno-ácido tetraacético tetraetil éster à base de Na + eléctrodo específico de iões espec�ico caracterizado por Messerli et. al. 22. Eletrodos íon-específicas também têm sido usados para medir concentrações de íons no líquido secretado por túbulos de Malpighi no ensaio Ramsay em 9,23 adulto e 16 larval Drosophila melanogaster, a Nova Zelândia Alpine Weta (Hemideina maori) 24 e 17 em mosquitos.
Aqui, descrevemos em detalhes o uso do Ramsay comodizer para medir as taxas de secreção de fluidos em túbulos de Malpighi de Drosophila melanogaster, bem como o uso de eletrodos de íon-específico para determinar as concentrações de K + e Na + dentro do fluido secretado e, portanto, o cálculo dos fluxos de iões transepithelial. Uma visão geral do ensaio é fornecida na Figura 1.
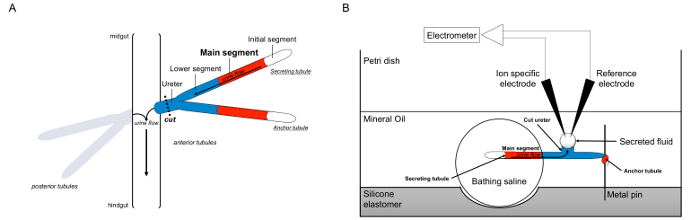
Figura 1. Esquema do Malpighian Palhinha eo Ramsay Ensaio com uso de íon-específicas eletrodos para medir concentrações de iões. Esta figura ilustra a configuração para o ensaio Ramsay. (A) Cada mosca tem quatro túbulos, um par de túbulos anterior e um par de túbulos posteriores, que flutuam na cavidade abdominal cercado por hemolinfa. Em cada par, as duas túbulos juntar-se no ureter, que, em seguida, esvazia-se a urina na junção do intestino médio e hindgut. Os túbulos são cegos-ended. A urina é gerado pelo segmento principal secretor de fluido (representado em vermelho), e flui para fora do ureter e no intestino. Após dissecção, a par dos túbulos é dissociada do intestino, ao seccionar o ureter. (B) A par dos túbulos é então transferida para uma gota de solução salina de banho dentro de uma cavidade do prato de ensaio. Um dos dois túbulos, referido aqui como o "túbulo âncora", é enrolado em torno de um pino de metal e é inerte. A outra é o túbulo túbulo secretora. O segmento inicial (que não secretam fluido) e segmento principal do túbulo segregam permanecer dentro da gotícula de solução salina de banho. Íons e movimento de água da salina de banho e para o lúmen do túbulo do segmento principal, em seguida, avançar em direção ao ureter, como ocorreria in vivo. O segmento inferior (azul) está fora da salina de banho e, portanto, inertes. Uma vez que o ureter é cortado, o fluido segregado surge como uma gota a partir da extremidade de corte do ureter. Tele secretada gota de fluido aumenta ao longo do tempo como a secreção continua, e o seu diâmetro é medido usando um micrómetro ocular. Uma camada de óleo mineral evita a evaporação do fluido que é segregado. Os eletrodos de referência específicas e de iões de medir a concentração de íons do fluido secretado. Por favor clique aqui para ver uma versão maior desta figura.
