T-lymfocytter er en gren af det adaptive immunsystem kendetegnet ved evnen til effektivt at genkende peptidantigen (Ag) bundet til major histocompatibility complex (MHC) molekyler gennem deres T-cellereceptorer (TCR'er) 1. Naive lymfocytter konstitutivt migrere og scanne faglige Ag celler '(APC'er; f.eks dendritiske celler) inden lymfeknuder, mens hukommelsen / effektor T-celler har brug for effektivt at overskue en meget bred vifte af APC og potentielle målceller inden perifere væv.
I min efter første indregning af beslægtet Ag på en APC, lymfocytter arrestere deres migration og begynder at danne en specialiseret intim celle-celle-interface betegnes "immunologisk synapse" (IS). Vedvarende (dvs. 30-60 min) kontakter er forpligtet til at forstærke og fastholde signalering 2-7 IS. Nye undersøgelser identificere, at inden for IS, er den kontinuerlige dannelse og hurtig remodeling af diskrete sub-cellulær signalering mikro-klynger (dvs. indeholdende MHC / Ag-TCR, F-actin, vedhæftning og signalmolekyler), der bestemmer styrken og kvaliteten af resulterende immunreaktioner 2-7. Dog er dynamiske detaljer og reguleringsmekanisme af denne proces ufuldstændigt forstået 8,9. Dette skyldes i høj grad fra de tekniske udfordringer forbundet med uregelmæssige topologier af APC overflader og dårligt kontrolleret orientering af celle-celle interaktion fly, spørgsmål, dybt begrænser den nødvendige Spatiotemporal billeddannelse nærmer 8-10 (Figure1A).
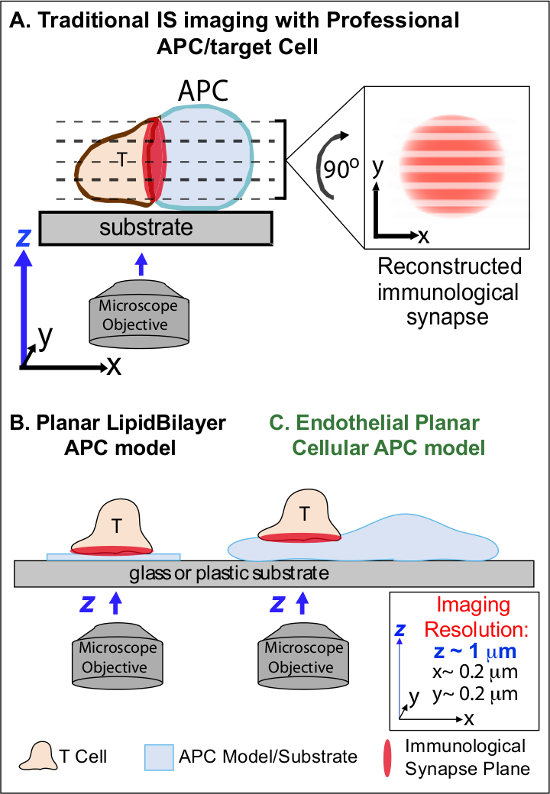
Figur 1. Et Fysiologisk Planar Cell APC Model for Imaging Immunologisk Synapse Dynamics. Den skematiske illustrerer traditionelle billeddannelse af immunologisk synapser mellem en T-celle og en professio udg APC (A) og T-celle og en traditionel plane lipiddobbeltlag APC model (B) i forhold til denne nye endotel plane APC model (C). Professionelle APC'er giver fysiologiske immunologiske synapser men tilbyder dårligt orienteret celle-celle-grænseflade (dvs. med hensyn til den optimale xy afbildningsplanet, opløsning ~ 0,2 um), som dramatisk kompromitterer rumlige (z billedplanet opløsning ~ 1 um) og tidsmæssige (dvs. på grund af behovet for gentagne gange at scanne gennem alle z imaging fly) opløsning på billeddannelse. Dobbeltlag modeller har en plan topologi, der giver optimal Spatiotemporal billedbehandling opløsning, men er også meget forenklet, ikke-fysiologiske og stive. Denne endotelcelle model kombinerer den plane topologi lipiddobbeltlag med fysiologisk substrat af en klassisk APC til at levere en optimal rumlig og tidsmæssig opløsning billeddannelse i et fysiologisk miljø.m / filer / ftp_upload / 53288 / 53288fig1large.jpg "target =" _ blank "> Klik her for at se en større version af dette tal.
Tidligere arbejde har delvist omgås disse hindringer ved at udvikle plane modeller (dvs. lipiddobbeltlag og antistof-coatede overflader), som giver optimal spatiotemporale opløsning (dvs. via fastsættelse af T-celleaktivering overflade i en enkelt plan, der er parallelt med den optimale xy imaging fly) 11-15 (figur 1B). Disse modeller har lettet vigtig indsigt i de subcellulære / molekylær dynamik, der styrer antigen signalering i T-celler, herunder opdagelsen af dynamiske actin / TCR signalering mikro-klynger 7,11-14. Imidlertid er sådanne modeller iboende oversimplificeres, samt stive (hinder udvikling / studie af 3-dimensionelle topologiske træk) (figur 1B). Derfor er det fortsat usikkert, hvordan at relatere sådanne resultater til grafisiologic celle-celle immunovervågning.
Men stadig dublerede er vaskulære og lymfatiske endotelceller fremstår som en stor (dvs. større i antal end alle professionelle APC'er, ved ~ 1000 gange) perifere rum af 'semi-professionelle "APC'er 16-18. Disse celler udtrykker MHC-I-, II- MHC-og et væld af co-stimulator molekyler (f.eks CD40, LFA3, ICOSL, 4-1 BB, OX40L, TL1A, PD-L1, men ikke CD80 og CD86), og er strategisk placeret ved blod-vævsgrænsefladen hvor de tjener specialiserede sentinel funktioner 16-18. Tidligere undersøgelser viste, at endotelceller effektivt kan re-stimulere effektor / hukommelse, men ikke naive, T-celler 19-25. Således spiller sandsynligvis unikke APC roller i effektorfasen af adaptive immunresponser inden for de perifere væv, såsom lokal indflydelse på T-celleaktivering, differentiering, hukommelse og tolerance 16,17,26 endotelceller. Critisk, når der dyrkes in vitro, endotelceller danner næsten plane celleoverflader og er let transficerbare (f.eks med fluorescerende protein reportere). Disse funktioner er ideelle til høj Spatiotemporal opløsning billeddannelse af topologiske dynamik under celle-celle interaktioner 19,27. Således endotelceller kunne tjene som en fysiologisk 'plane cellulære APC' model udpræget egnet til undersøgelse af de subcellulære / molekylære mekanismer remodeling som driver antigen-genkendelse og regulerer responser (figur 1C) 19,20.
Tidligere konstaterede komplementære billeddannelsesteknikker (herunder transfektion af endotel celler med fluorescerende protein skaberne af plasmamembranen og cytosol) for at studere detaljerne i leukocyt-endotel interaktion under adhæsion og transendotelial migration 27, viste, at leukocytter aktivt sonde overfladen af endotel ved dynamisk indsættelse af end tilbagetrækning af sub-micron-skala, actin-rige cylindriske fremspring (200-1000 nm i diameter og dybde ~) betegnes invadosome-lignende fremspring (dvs. "ILPS ') 27,28. Disse billeddannende fremgangsmåder er blevet udvidet yderligere sammen med oprettelsen af protokoller for at drage fordel af endotel APC-funktion for at udvikle de første metoder til høj spatiotemporale opløsning billeddannelse af T-celle-endothelial immunologisk synapse som rapporteret 19,20 og beskriver endvidere heri. En central konklusion stammer fra denne roman plane cellulære APC-modellen er, at T-celle efterværn fungere både i at fremme indledende Ag detektion og fastholde efterfølgende signalering. Faktisk arrays af flere efterværn (der blev stabiliseret og påløbne som reaktion på indledende calciumflux) viser berigelse i TCR og molekyler, der tyder på aktiv signalering sådan PKC-Q, ZAP-70, phosphotyrosin og HS1. Derfor synes efterværn at repræsentere et tredimensionalt fysiologisk svarer til TCR-signalering mikroklynger set i plane dobbeltlag modeller. Denne tilgang, således følsomt afslører / rapporter molekylære og arkitektoniske (og underforståede biomekaniske) dynamik ellers ikke påvises.
Den her beskrevne metode bør være nyttige for at bygge bro mellem professionelle APC og kunstige APC substrat modeller for at forbedre vores evne til at afhøre grundlæggende mekanismer i adaptive immunrespons. Mens her er fokus på aktiveringen af CD4 + Th1-typen effektor / lagercelle, kan denne grundlæggende fremgangsmåde let modificeres til at studere en bred vifte af T-celle typer og Ags, som beskrevet nedenfor.
