Os linfócitos T são um ramo do sistema imunitário adaptativo, caracterizado por a capacidade de reconhecer eficientemente antigénio peptídico (Ag) ligado ao complexo principal de histocompatibilidade (MHC) através dos seus receptores de células T (TCRs) 1. Linfócitos naive constitutivamente migrar e digitalizar 'Ag células apresentadoras de profissionais "(APCs; por exemplo, células dendríticas) dentro de gânglios linfáticos, enquanto as células de memória / T efetoras precisa fazer um levantamento efetivamente uma gama extremamente ampla de APCs e células alvo em potencial dentro tecidos periféricos.
Na min após o reconhecimento inicial de cognato Ag em um APC, linfócitos prender sua migração e começam a formar uma interface célula-célula íntimo especializado denominado "sinapse imunológica" (IS). Sustentada (ie, 30-60 min) É contatos são necessários para ampliar e sustentar a sinalização 2-7. Estudos emergentes identificar que no seja, é a formação contínua e rápida remodeling de micro-aglomerados discretos sinalização sub-celular (ou seja, contendo MHC / Ag-TCR, F-actina, adesão e moléculas de sinalização) que determinam a força e qualidade dos resultando respostas imunes 2-7. No entanto, detalhes dinâmicos e mecanismo de regulação deste processo são completamente compreendidas 8,9. Isso decorre, em grande parte dos desafios técnicos associados com topologias de superfícies irregulares APC e orientação mal controlada dos planos de interação célula-célula, as questões que limitam profundamente a imagem espaço-temporal requisito aproxima 8-10 (Figura 1A).
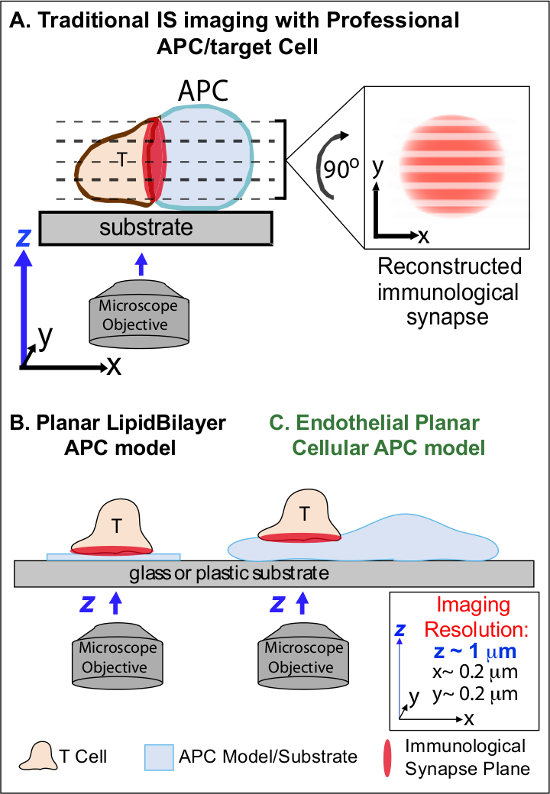
Figura 1. Um Fisiológica Planar Modelo APC celular for Imaging imunológica Synapse Dynamics. O esquema ilustra imagem tradicional da sinapse imunológica entre uma célula T e um professio nal APC (A) e de células T e um lipídicas planas modelo tradicional bicamada APC (B) em comparação com este modelo planar APC endotelial novel (C). APCs profissionais fornecer sinapses imunológicos fisiológicos, mas oferecem interface de mal orientadas célula-célula (isto é, em relação ao plano de imagem XY óptima; resolução ~ 0,2 ^ M), o que compromete dramaticamente espacial (plano de imagem z resolução ~ 1 mm) e temporal (isto é, devido à necessidade de digitalizar repetidamente através de todos os planos de imagem z) resolução de imagem. Modelos de duas camadas tem uma topologia plana que fornece melhor resolução de imagem espaço-temporal, mas também são altamente simplificado, não-fisiológica e rígida. Este modelo combina células endoteliais a topologia planar de camadas duplas lipídicas com o substrato fisiológico de um APC clássico para oferecer resolução de imagem espacial e temporal ideal em um ambiente fisiológico.m / files / ftp_upload / 53288 / "target =" _ blank 53288fig1large.jpg "> Clique aqui para ver uma versão maior desta figura.
Trabalhos anteriores contornada parcialmente estes obstáculos através do desenvolvimento de modelos de substrato planar (isto é, as bicamadas lipídicas e das superfícies revestidas com anticorpo) que fornecem resolução espaço-temporal óptimo (ou seja, através de fixação superfície a activação das células T num único plano que é paralelo ao imagiologia XY óptima avião) 11-15 (Figura 1B). Estes modelos têm facilitado importantes insights sobre a dinâmica subcelulares / molecular que controlam a sinalização antigênico nas células T, incluindo a descoberta de actina dinâmica / TCR sinalização micro-aglomerados 7,11-14. No entanto, tais modelos são inerentemente muito simplificado, assim como rígida (impedindo o desenvolvimento / estudo de características topológicas 3-dimensional) (Figura 1B). Portanto, permanece incerto como relacionar esses achados para phycélula-célula siologic vigilância imunológica.
Embora ainda pouco estudadas, vascular e células endoteliais linfáticas estão emergindo como uma grande (ou seja, maior em número do que todos os APCs profissionais, por ~ 1.000 vezes) compartimento periférico de "semi-profissional" APCs 16-18. Estas células expressam MHC-I, MHC-II e uma multiplicidade de moléculas co-estimuladoras (por exemplo, CD40, LFA3, ICOSL, 4-1BB, OX40L, TL1A, PD-L1, mas não CD80 e CD86) e são estrategicamente posicionada na interface de sangue-tecido onde servem funções especializadas sentinela 16-18. Estudos anteriores demonstraram que as células endoteliais podem efetivamente re-estimular efector / memória, mas não naïve, células T 19-25. Assim, as células endoteliais são susceptíveis de desempenhar papéis exclusivos da APC na fase efectora da resposta imune adaptativa no interior dos tecidos periféricos, tais como a influência local sobre a activação de células T, a diferenciação, a memória e a tolerância 16,17,26. Criticalmente, quando cultivadas in vitro, as células endoteliais formam superfícies celulares praticamente planares e estão facilmente transfectáveis (por exemplo, com os repórteres de proteína fluorescente). Estas características são ideais para imagens de alta resolução espacial e temporal da dinâmica topológica durante as interações célula-célula 19,27. Assim, as células endoteliais pode servir como um fisiológico "planar celular da APC modelo extremamente adequado para o estudo dos mecanismos de remodelação subcelulares / moleculares que conduzem o reconhecimento do antigénio e regulam as respostas (Figura 1C) 19,20.
Previamente estabelecida técnicas de imagem complementares (incluindo a transfecção de células endoteliais com fabricantes de proteína fluorescente da membrana de plasma e citossol) para estudar os detalhes da interacção de leucócitos-endotélio durante a adesão e a migração transendotelial de 27, mostraram que os leucócitos sondar activamente a superfície do endotélio por dinâmico inserção de umd retração de sub-mícron-escala, saliências cilíndricas ricas em actina (~ 200-1.000 nm de diâmetro e profundidade) denominado saliências invadosome-like (ou seja, 'ILPS') 27,28. Estas abordagens de imagem têm sido expandida, juntamente com a criação de protocolos para tirar proveito da função endotelial APC para desenvolver os primeiros métodos para imagens de alta resolução espacial e temporal da célula endotelial sinapse imunológica T como relatado 19,20 e ainda descrever aqui. Uma constatação fundamental derivado dessa modelo APC celular romance planar é que procedimentos ILP células T funcionar tanto na promoção da detecção Ag inicial e na manutenção de sinalização subsequente. Na verdade, matrizes de múltiplos procedimentos ILP (que foram estabilizados e acumulados em resposta a rubricar o fluxo de cálcio) mostram enriquecimento em TCR e moléculas de sinalização sugestivos activa, tais PKC-Q, ZAP-70, fosfotirosina e HS1. Portanto, procedimentos ILP parecem representar um equivalente fisiológico tridimensional para o micro-TCR sinalizaçãoaglomerados observados nos modelos de duas camadas planas. Esta abordagem, assim, com sensibilidade revela / relatórios dinâmica molecular e arquitetônicos (e implícitas biomecânico) de outra forma não detectável.
O método descrito aqui deve ser útil para fazer a ponte entre APC profissional e modelos artificiais de substrato APC a fim de aumentar a nossa capacidade de interrogar mecanismos básicos da resposta imune adaptativa. Embora aqui o foco é sobre a activação das células T CD4 + do tipo Th1 efector / célula de memória, esta abordagem básica pode ser facilmente modificado para estudar uma ampla gama de tipos de células T e Ags, como discutido abaixo.
