T lenfositleri majör histokompatibilite kompleksi (MHC) bağlı verimli peptit antijeni (Ag) tanıma yeteneği ile karakterize adaptif bağışıklık sisteminin bir dalı vardır, T hücre reseptörlerinin (TCR'ler) 1 ile molekülleri. Naif lenfositler kurucu göç ve profesyonel Ag sunan hücreler '(ZPT; örneğin, dendritik hücreler) tarama bellek / efektör T hücreleri etkin bir periferik dokularda APC' ve potansiyel hedef hücrelerin son derece geniş bir yelpazede anket gerekir iken, lenf düğümleri içinde.
Bir APC üzerindeki soydaş Ag ilk muhasebeleştirilmesi aşağıdaki min, bunların göç tutuklama ve özel bir samimi hücre-hücre arayüzü olarak adlandırılan 'immünolojik sinaps' oluşmaya başlar lenfosit (IS). Sürekli (yani 30-60 dk) temas yükseltmek ve 2-7 sinyalizasyon sürdürmek için gerekli olan IS. Gelişmekte olan çalışmalar IS içinde, sürekli oluşumu ve hızlı r olduğunu tespitgücü ve immün yanıtları 2-7 sonuçlanan kalitesini belirlemek (örneğin, MHC / Ag-TCR, F-aktin, yapışma ve sinyal molekülleri içeren) ayrık alt hücresel sinyal mikro-kümelerin emodeling. Ancak, bu sürecin dinamik detaylar ve düzenleyici mekanizma tam olarak 8,9 anlaşılmaktadır. Bu son derece gerekli zamanmekansal görüntüleme sınırı APC yüzeylerin düzensiz topolojileri ve hücre-hücre etkileşimi uçakları kötü kontrollü yönlendirme, sorunları ile ilgili teknik zorlukları büyük ölçüde kaynaklanmaktadır 8-10 (Figure1A) yaklaşır.
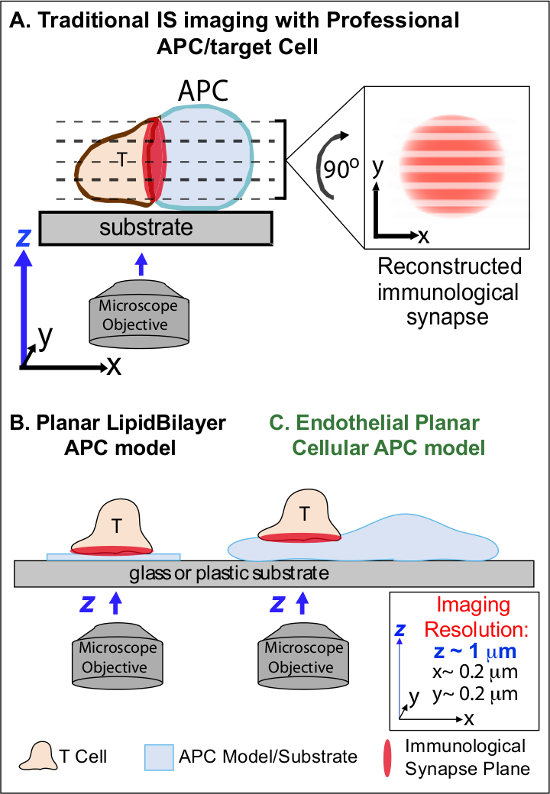
Görüntüleme İmmünoloji Synapse Dynamics Şekil 1. Fizyolojik Planar hücre APC Modeli. Şematik bir T hücresi ve professio arasındaki immünolojik sinaps geleneksel görüntüleme göstermektedir nal APC (A) ve T hücresi ve bu yeni endotel düzlemsel APC modelinde (C) ile karşılaştırıldığında, geleneksel bir düzlemsel lipid iki katmanlı APC modeli (B). , Dramatik, yani mekansal (z görüntüleme düzey çözünürlük ~ 1 mikron) ve temporal (ödün; Profesyonel ZPT fizyolojik immünolojik sinaps sağlar, ancak (çözünürlük ~ 0.2 mikron Optimum xy görüntüleme düzlemine göre yani) kötü amaçlı hücre-hücre arabirimi sunar ihtiyacı nedeniyle defalarca görüntüleme çözünürlüğü) tüm z görüntüleme uçakları ile tarayın. Iki tabakalı modelleri Optimal zamanmekansal görüntüleme çözünürlüğü sağlar düzlemsel topoloji var, ama aynı zamanda son derece, fizyolojik olmayan ve sert basitleştirilmiştir. Bu endotel hücre modeli fizyolojik bir ortamda optimum mekansal ve zamansal görüntüleme çözünürlüğü sağlamak için klasik bir APC fizyolojik substrat ile lipit bilayers düzlemsel topolojisini birleştirir.m / files / ftp_upload / 53288 / 53288fig1large.jpg "target =" _ blank "> bu rakamın büyük halini görmek için lütfen buraya tıklayınız.
Önceki çalışma kısmen uygun xy görüntüleme paralel olan tek bir planına T hücresi aktivasyonu aracılığı ile yüzey sabitleme örneğin, uygun uzaysal çözünürlük (sağlar geliştirmek düzlemsel alt-tabaka bir model (yani, lipid çift tabakaları ve antikor-kaplı yüzeyler), bu engeller etrafından olan düzlem) 11-15 (Şekil 1B). Bu modeller, dinamik aktin / TCR sinyalizasyon mikro kümelerin 7,11-14 keşif dahil olmak üzere T hücrelerinin antijenik sinyalizasyon, kontrol moleküler / hücre içi dinamikleri önemli bilgiler kolaylaştırmıştır. Bununla birlikte, bu modeller doğal basitleştirilmiş edilir, yanı sıra, katı (Şekil 1B) (3-boyutlu bir topolojik özelliklerin geliştirilmesi / çalışma engelleyen). Bu nedenle, phy böyle bulguları ilişkilendirmek nasıl belirsizdirsiologic hücre-hücre immün gözetim.
Hala understudied rağmen, damar ve lenfatik endotel hücreleri büyük olarak ortaya çıkmaktadır (yani ~ 1000 kat tüm profesyonel APC'lerce daha numaraları, daha fazla) 'yarı profesyonel' APC '16-18 periferik bölmesi. (Örneğin, CD40, LFA3, ICOSL, 4-1BB, OX40L, TL1A, PD-L1; fakat CD80 ve CD86) Bu hücreler, MHC-I-, MHC-II-ve ko-stimülatör moleküllerin çok sayıda dile ve stratejik olarak Onlar özel nöbetçi işlevleri 16-18 hizmet kan doku arayüzünde konumlandırılmış. Daha önceki çalışmalar, endotel hücrelerinin etkili bir efektör / bellek ama saf olmayan, T hücrelerini 19-25 yeniden stimüle edebilirler. Bu nedenle, endotelyal hücreler gibi T hücresi aktivasyonu, farklılaşması, hafıza ve tolerans 16,17,26 yerel etkisi gibi periferik dokular içinde adaptif bağışıklık tepkilerinin, efektör aşamasında özel APC rol oynaması olasıdır. Criin vitro yetiştirilen ola- rak, endotel hücreleri neredeyse düzlemsel hücre yüzeylerine formu (floresan protein muhabir olan, örneğin) kolayca transfekte bulunmaktadır. Bu özellikler, hücre-hücre etkileşimleri 19,27 sırasında topolojik dinamiklerin yüksek uzaysal çözünürlüklü görüntüleme için idealdir. Böylece endotel hücreleri tepkileri (Şekil 1C) 19,20 antijen tanıma sürücü ve düzenleyen moleküler / hücre içi biçimlenme mekanizmalarının çalışma için belirgin uygun bir fizyolojik 'düzlemsel hücresel APC' model olarak hizmet olabilir.
Daha önce yapışma ve transendoteliyal göç 27 sırasında lökosit-endotel etkileşimi detaylarını incelemek için (plazma zarı ve sitoplazmada floresan protein üreticileri ile endotelin hücrelerin transfeksiyonu dahil) tamamlayıcı görüntüleme tekniklerini kurulan lökositler aktif, dinamik tarafından endotel yüzeyine soruşturma olduğunu gösterdi Ekleme birmikronaltı ölçek d retraksiyonu, aktin-zengin silindirik çıkıntılar (~ çap ve derinlikte 200-1,000 nm) invadosome benzeri çıkıntılar (yani, 'ILPS') 27,28 olarak nitelendirdi. Bu görüntüleme yaklaşımları daha da bildirilen 19,20 olarak T hücre-endotelyal immünolojik sinaps yüksek uzaysal çözünürlüklü görüntüleme için ilk yöntemler geliştirmek endotel APC fonksiyonunun yararlanmak ve ayrıca burada açıklamak için protokollerin oluşturulması ile birlikte genişletilmiştir. Bu yeni düzlemsel hücresel APC modelinden elde edilen bir merkez bulgu T hücresi ILPS ilk Ag algılama teşvik ve sonraki sinyalizasyon sürdürülmesi hem işlev olmasıdır. Nitekim, (stabilize ve kalsiyum akı başlangıç yanıt tahakkuk eden) TCR ve aktif sinyal gibi PKC-Q, ZAP-70, fosfotirosinin ve HS1 düşündüren moleküllerde gösteri zenginleştirme birden ILPS'nin diziler. Bu nedenle, ILPS TCR-sinyal mikro üç boyutlu fizyolojik eşdeğer temsil etmek gibi görünüyordüzlemsel iki tabakalı modellerinde görülen kümeler. Bu yaklaşım, bu nedenle, hassas ortaya / raporlar, moleküler ve mimari (ve biyomekanik zımni) dinamikleri aksi saptanabilir değil.
Burada açıklanan yöntem adaptif bağışıklık tepkilerinin temel mekanizmaları sorgulamak için yeteneğimizi geliştirmek amacıyla profesyonel APC ve yapay APC substrat modelleri arasında köprü için yararlı olacaktır. Burada odaklama CD4 + efektör Th1-tipi / hafıza hücresinin aktivasyonuna iken, aşağıda ele alındığı gibi, bu temel yaklaşım, hali hazırda, T hücre türleri ve Ag'lerin geniş bir çalışma için modifiye edilebilir.
