T-lymfocytter er en gren av det adaptive immunsystemet, karakterisert ved evnen til effektivt å gjenkjenne peptid antigen (Ag) bundet til hovedhistokompatibilitetskompleks (MHC) molekyler gjennom deres T-celle-reseptorer (TCR) 1. Naive lymfocytter konstitutivt migrere og skanne faglige Ag celler '(APC, for eksempel, dendrittiske celler) i lymfeknuter, mens minne / effektor T-celler trenger for å effektivt kartlegge et ekstremt bredt spekter av APC og potensielle målceller i perifere vev.
I min etter første gangs registrering av beslektet Ag på en APC, lymfocytter arrestere deres migrasjon og begynner å danne en spesialisert intim celle-celle-grensesnitt kalles 'immunologiske synapsen "(IS). Vedvarende (dvs. 30-60 min) ER kontakter er nødvendig for å forsterke og opprettholde signale 2-7. Emerging studier identifisere at innenfor IS, er det kontinuerlig dannelse og rask remodeling av diskrete sub-cellulære signalmikro klynger (dvs. inneholder MHC / Ag-TCR, F-aktin, heft og signalmolekyler) som bestemmer styrken og kvaliteten på resulterende immunresponser 2-7. Imidlertid er dynamiske detaljer og reguleringsmekanisme av denne prosessen ufullstendig forstått 8,9. Dette stammer i stor grad fra tekniske utfordringer knyttet til uregelmessige topologier av APC flater og dårlig kontrollert orientering av celle-celle interaksjons fly, problemer som dypt begrenser den nødvendige tid og rom bildebehandling nærmer 8-10 (Figure1A).
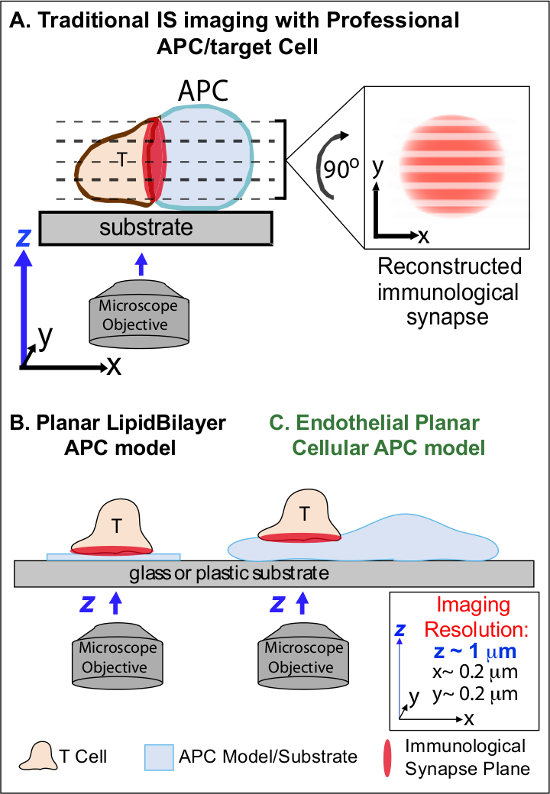
Figur 1. En fysiologisk Planar Cell APC Modell for Imaging Immunologisk Synapse Dynamics. Den skjematisk illustrerer tradisjonelle avbildning av immunologisk synapse mellom en T-celle og en professio nal APC (A) og T-celle og en tradisjonell plan lipidbilag APC modellen (B) i forhold til denne nye endotelial plane APC-modellen (C). Profesjonelle APC tilveiebringe fysiologiske immunologiske synapser men har dårlig orientert celle-celle grensesnitt (dvs. med hensyn til den optimale xy avbildningsplanet; oppløsning ~ 0,2 mikrometer), noe som dramatisk kompromisser romlig (z avbildningsplanet oppløsning ~ 1 um) og tidsmessig (dvs. på grunn av behovet for å gjentatte ganger søke gjennom alle z avbildningsplan) oppløsning av bildebehandling. Bilags modeller har en plan topologi som gir optimal spatiotemporal bildeoppløsning, men er også svært forenklet, ikke-fysiologiske og rigid. Dette endotelceller modellen kombinerer planar topologien lipidbilag med fysiologisk substrat av en klassisk APC å levere optimal romlig og tidsmessig bildeoppløsning i en fysiologisk setting.m / filer / ftp_upload / 53288 / 53288fig1large.jpg "target =" _ blank "> Klikk her for å se en større versjon av dette tallet.
Tidligere arbeid har delvis omgå disse hindringene ved å utvikle plant substrat modeller (dvs. lipidbilag og antistoff-belagte overflater) som gir optimal tid og rom-oppløsning (dvs. via fiksering av T-celleaktivering overflaten til et enkelt plan som er parallelt til den optimale xy avbildnings planet) 11-15 (figur 1B). Disse modellene har tilrettelagt viktig innsikt i de subcellulære / molekylær dynamikk som styrer antigen signale i T-celler, inkludert oppdagelsen av dynamisk aktin / TCR signal mikro-klynger 7,11-14. Imidlertid er slike modeller iboende kles, samt stiv (utelukker utviklingen / studiet av tre-dimensjonale topologiske egenskaper) (figur 1B). Derfor er det fortsatt usikkert hvordan å fortelle om slike funn til grafisiologic celle-celle immunovervåkning.
Men fortsatt studerte, er vaskulære og lymfatiske endotelceller fremstår som en stor (dvs. større i antall enn alle profesjonelle APC, ved ~ 1000 ganger) perifer rommet i "semi-profesjonelle 'APC 16-18. Disse cellene uttrykker MHC-I-, MHC-II- og et mangfold av co-stimulatorer molekyler (f.eks CD40, LFA3, ICOSL, 4-1BB, OX40L, TL1A, PD-L1, men ikke CD80 og CD86) og er strategisk plassert på blod-vev grensesnitt der de tjener spesialiserte sentinel funksjoner 16-18. Tidligere studier har vist at endotelcellene effektivt kan gjen stimulere effektor / minne, men ikke naive, T-celler 19-25. Således endotelceller vil kunne spille unike APC roller i effektorfasen adaptive immunresponser i de perifere vev, slik som lokale innflytelse på T-celleaktivering, differensiering, hukommelse og toleranse 16,17,26. Critisk, når dyrket in vitro, endotelceller danner praktisk talt plane celleoverflater og er lett transfectable (f.eks, med fluorescerende protein reportere). Disse funksjonene er ideelle for høy spatiotemporal oppløsning avbildning av topologiske dynamikk i celle-celle interaksjoner 19,27. Dermed endotelceller kan tjene som en fysiologisk 'planar cellular APC' modell utpreget egnet for studiet av subcellulære / molekylær ombygging mekanismer som driver antigen anerkjennelse og regulerer svar (figur 1C) 19,20.
Tidligere etablert komplementære billedteknikker (inkludert transfeksjon av endothelia cellene med fluorescente protein produsentene av plasmamembranen og cytosol) for å studere detaljene for leukocytt-endotelial interaksjon under adhesjon og transendothelial migrering 27, viste at leukocytter sonde aktivt overflaten av endotel ved dynamisk innsetting end tilbaketrekking av sub-mikron-skala, actin-rik sylindriske fremspring (~ 200-1000 nm i diameter og dybde) betegnes invadosome lignende fremspring (dvs. 'ILPs') 27,28. Disse avbildnings tilnærminger har blitt ytterligere utvidet sammen med etableringen av protokoller for å dra nytte av endotelial APC-funksjon til å utvikle det første metoder for høy spatiotemporal oppløsning avbildning av T-celle-endotelial immunologiske synapse som rapportert 19,20 og beskriver videre heri. Et sentralt funn som stammer fra denne romanen planar mobil APC-modellen er at T-celle ILPs fungere både i å fremme innledende Ag deteksjon og i å opprettholde påfølgende signalering. Faktisk, matriser av flere ILPs (som ble stabilisert og påløpt i respons til innledende kalsium flux) viser berikelse i TCR og molekyler som tyder på aktiv signal slik PKC-Q, ZAP-70, fosfotyrosin og HS1. Derfor ILPs synes å representere en tredimensjonal fysiologisk ekvivalent med den TCR-signalisering mikroklynger sett i planar bilags modeller. Denne tilnærmingen, og dermed avslører følsomt / rapporter molekylære og arkitektoniske (og implisitte biomekaniske) dynamikk ellers ikke synlig.
Metoden er beskrevet i dette dokumentet skal være nyttig for å bygge bro over gapet mellom profesjonell APC og kunstige APC substrat modeller for å styrke vår evne til å avhøre grunnleggende mekanismer for adaptive immunresponser. Mens her gjelder det på aktivering av CD4 + Th1-type effektor / minnecelle, kan denne grunnleggende metode kan lett modifiseres for å studere et bredt spekter av T-celletyper og Ags, som diskutert nedenfor.
