T-lymfocyter är en gren av det adaptiva immunsystemet kännetecknas av förmågan att effektivt känna igen peptidantigen (Ag) bundna till större histokompatibilitetskomplex (MHC) molekyler genom deras T-cellreceptorer (TCR) 1. Naiva lymfocyter konstitutivt migrera och skanna yrkesmässiga Ag presenterande celler "(APC, t.ex. dendritiska celler) i lymfkörtlar, medan minnet / T-celler behöver för att effektivt kartlägga ett mycket brett utbud av APC och potentiella målceller inom perifera vävnader.
I min efter första redovisningen av besläktad Ag på en APC, lymfocyter arrestera deras migration och börja bilda en specialiserad intim cell-cell-gränssnittet kallas "immunologisk synaps" (IS). Fortsatt (dvs 30-60 min) ÄR kontakter krävs för att förstärka och upprätthålla signalering 2-7. Tillväxtstudier identifiera att inom IS, är det den kontinuerliga bildningen och snabb remodeling diskreta sub-cellulära signaleringsmikrokluster (dvs innehållande MHC / Ag-TCR, F-aktin, vidhäftning och signalmolekyler) som avgör styrka och kvalitet resulterande immunsvar 2-7. Men dynamiska detaljer och regleringsmekanism i denna process ofullständigt förstått 8,9. Detta beror till stor del från tekniska utmaningar som är förknippade med oregelbundna topologier av APC ytor och dåligt kontrollerad orientering av interaktions cell-cell flygplan, frågor som i grunden begränsar den erforderliga Spatiotemporal avbildning närmar 8-10 (Figure1A).
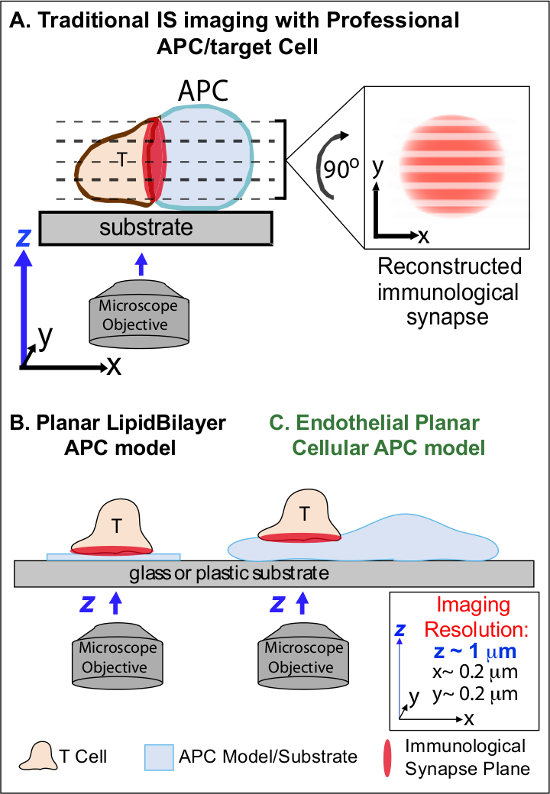
Figur 1. en fysiologisk Planar Cell APC Modell för Imaging Immunological Synapse Dynamics. Schemat visar traditionella avbildning av immunologisk synaps mellan en T-cell och en professio nella APC (A) och T-cell och en traditionell plana lipiddubbelskikt APC modell (B) i jämförelse med denna nya endotel plana APC modell (C). Professionella APC ger fysiologiska immunologiska synapser men erbjuder dåligt orienterad cell-cell-gränssnitt (det vill säga, med avseende på den optimala xy avbildningsplanet; upplösning ~ 0,2 | im), som dramatiskt äventyrar rumsliga (z avbildningsplanet upplösning ~ 1 | im) och tidsmässiga (dvs., på grund av behovet att upprepade gånger söka igenom alla z avbildningsplan) upplösning avbildning. Dubbelskikts modeller har ett plant topologi som ger optimal Spatiotemporal avbildningsupplösning, men är också mycket förenklade, icke-fysiologiska och styvt. Denna endotelceller modellen kombinerar den plana topologi lipidbiskikt med den fysiologiska substrat av ett klassiskt APC att leverera optimal rumsliga och tidsmässiga avbildning upplösning i en fysiologisk miljö.m / filer / ftp_upload / 53288 / 53288fig1large.jpg "target =" _ blank "> Klicka här för att se en större version av denna siffra.
Tidigare arbete har delvis kring dessa hinder genom utvecklings plana substratmodeller (dvs lipiddubbelskikt och antikroppsbelagda ytor) som ger optimala Spatiotemporal upplösning (dvs., via fastställande av cellaktivering ytan T till en och samma plan som är parallellt med den optimala xy avbildning planet) 11-15 (Figur 1B). Dessa modeller har underlättat viktiga insikter i de subcellulära / molekylära dynamik som styr antigen signalering i T-celler, inklusive upptäckten av dynamiska aktin / TCR signalering mikrokluster 7,11-14. Men sådana modeller inneboende förenklad, liksom stela (utgör hinder för utveckling / studie av 3-dimensionella topologiska funktioner) (Figur 1B). Därför är det osäkert hur man relatera dessa resultat till physiologic cell-cell-immunövervakning.
Men fortfarande understudied är vaskulära och lymfatiska endotelceller framstår som en stor (dvs större i antal än alla professionella APC, genom ~ 1000 gånger) periferiutrymme "halvprofessionella" APC 16-18. Dessa celler uttrycker MHC-I-, MHC-II och en mängd co-stimulator molekyler (t.ex., CD40, LFA3, ICOSL, 4-1 BB, OX40L, TL1A, PD-L1, men inte CD80 och CD86) och är strategiskt placerad vid blodvävnads gränssnitt där de tjänar specialiserade kontrollfunktionerna 16-18. Tidigare studier har visat att endotelceller kan effektivt åter stimulera effektor / minne, men inte naiva, T-celler 19-25. Sålunda endotelceller är sannolikt att spela unika APC roller i effektorfasen av adaptiva immunresponser inom de perifera vävnader, såsom lokalt inflytande på T-cellsaktivering, differentiering, minne och tolerans 16,17,26. Critiskt, när den odlas in vitro, endotelceller bildar nästan plana cellytor och är lätt transfekterbara (t.ex. med fluorescerande protein reportrar). Dessa funktioner är idealiska för hög Spatiotemporal upplösning avbildning av topologiska dynamik under cell-cell-interaktioner 19,27. Således endotelceller kan tjäna som en fysiologisk "plana cell APC modell klart lämpad för studier av de subcellulära / molekylära remodeling mekanismer som driver antigenigenkänning och reglerar svar (Figur 1C) 19,20.
Tidigare etablerade kompletterande avbildningstekniker (inklusive transfektion av endotel celler med fluorescerande protein skaparna av plasmamembranet och cytosolen) för att studera detaljerna i leukocyt-endotel interaktion under vidhäftning och transendotelial migration 27, visade att leukocyter sond aktivt yta endotel genom dynamisk insättnings end indragning av sub-micron skala, aktin rika cylindriska utsprång (~ 200-1000 nm i diameter och djup) kallas invadosome liknande utsprång (dvs "IUP) 27,28. Dessa avbildning har utökats ytterligare tillsammans med skapandet av protokoll för att dra nytta av endotel APC-funktion för att utveckla de första metoderna för hög Spatiotemporal upplösning avbildning av T-cells endothelial immunologisk synaps som rapporterats 19,20 och ytterligare beskriver häri. En central slutsats härrör från denna nya plana cellulära APC-modellen är att T-cell IUP fungera både för att främja inledande Ag upptäckt och för att upprätthålla efterföljande signalering. I själva verket, kedjor av flera IUP (som stabiliserats och upplupna som svar på initial kalciumflöde) visar anrikning i TCR och molekyler som tyder på aktiv signalering sådan PKC-Q, ZAP-70, fosfotyrosin och HS1. Därför IUP verkar representera en tre-dimensionell fysiologisk motsvarande TCR-signalering mikrokluster sett i plana dubbelskiktsmodeller. Detta tillvägagångssätt således känsligt avslöjar / rapporter molekylära och arkitektoniska (och underförstådda biomekaniska) dynamik annars inte upptäckas.
Den metod som beskrivs i detta dokument ska vara till nytta för att överbrygga klyftan mellan professionella APC och konstgjorda APC substrat modeller för att förbättra vår förmåga att förhöra grundläggande mekanismer adaptiva immunsvar. Även här ligger fokus på aktivering av CD4 + Th1-typ effektor / minnescell, kan denna grundläggande strategi lätt modifieras för att studera ett brett spektrum av T-celltyper och Ags, såsom diskuteras nedan.
