The in vivo alkaline comet assay was performed in conjunction with the enzyme-modified comet assay to measure both direct and oxidative DNA damage in the liver of rats treated with cyproterone acetate (CPA)5. CPA is a synthetic hormonal drug that induces rat liver tumors in a sex-specific manner, with five-fold higher doses needed to induce liver tumors in male rats compared to females24. We found that the direct DNA damage produced by CPA in the liver of male and female rats has the same sex-specific pattern as its hepatotumorigenicity: a five-fold-higher dose of CPA is needed to induce a significant increase in DNA damage in the livers of males compared to females (Figure 1).
We hypothesize that the sex-specific comet assay result is due to the activity of hydroxysteroid sulfotransferase(s) (HST), which is 15-fold higher in adult female rats than in adult males. HST metabolism is a rate-limiting step in the activation of CPA to DNA-binding metabolite(s)25. In contrast, CPA-induced oxidative DNA damage was generally greater in male than female rat livers, and thus less likely to be a rate-limiting step in tumor formation (Figure 2). Histopathology evaluation of livers from CPA-treated rats showed no evidence of agent-induced apoptosis or necrosis (Tables 1 and 2), indicating that the positive comet assay results were not a secondary effect of cytotoxicity5. Figures 1 & 2 and Tables 1 & 2 are reprinted with permission of Elsevier B.V. (Reference 5).
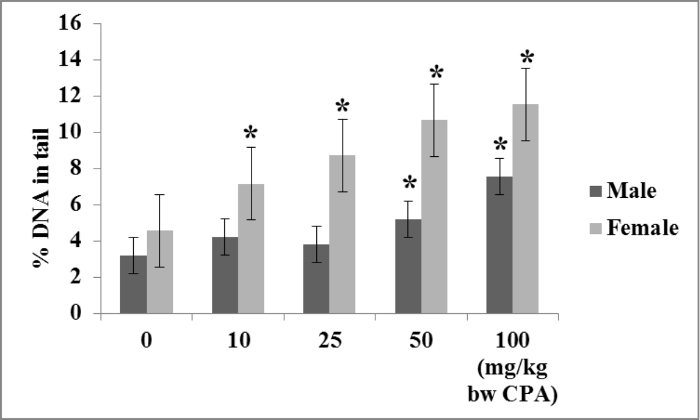
Figure 1. DNA damage in livers of CPA-treated male and female rats measured with the in vivo alkaline comet assay. Groups of five seven-week-old male and female F344 rats were treated with olive oil or with 10, 25, 50, or 100 mg/kg/day CPA in olive oil. Treatments were conducted at 0, 24, and 45 hr, the rats were sacrificed at 48 hr, and DNA damage was measured in liver as % tail DNA using the alkaline comet assay. CPA treatment induced an increase in % tail DNA in the livers of male rats in a threshold-like manner, with significant increases being detected only with the 50 and 100 mg/kg/day doses. CPA treatment induced DNA strand breaks in the livers of female rats in a near-linear dose-dependent manner, with significant increases in % tail DNA detected in all CPA groups. The Lowest Observed Genotoxicity Effect Levels (LOGELs) for CPA-induced DNA damage in the livers of female and male rats were estimated to be 10 and 50 mg/kg/day, respectively. *Significant at p≤0.05 relative to the vehicle control; error bars represent standard deviation. Please click here to view a larger version of this figure.
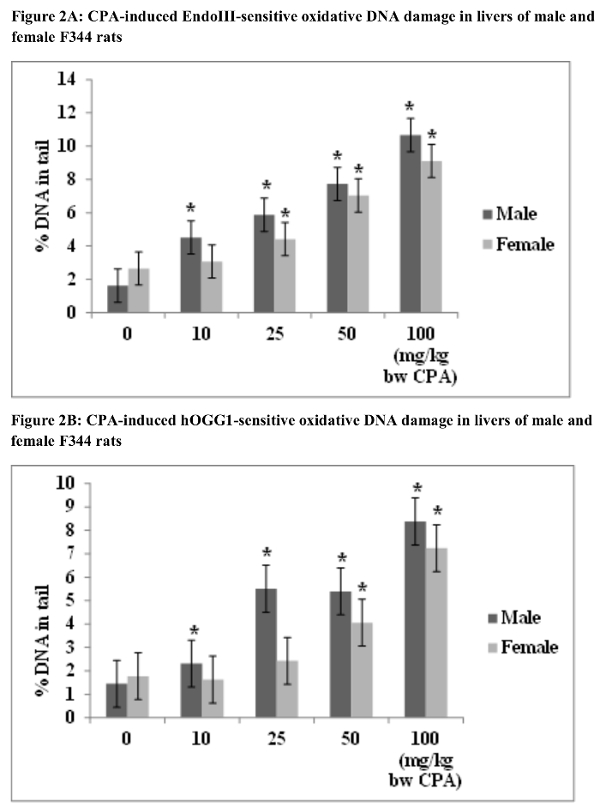
Figure 2. Oxidative DNA damage in livers of CPA-treated male and female rats measured with the enzyme-modified in vivo alkaline comet assay. Groups of five seven-week-old male and female F344 rats were treated with olive oil or with 10, 25, 50, or 100 mg/kg/day CPA in olive oil. Treatments were conducted at 0, 24, and 45 hr, the animals were sacrificed at 48 hr, and the enzyme-modified comet assay was conducted using % tail DNA as a metric of DNA damage. All CPA doses produced significant increases in Endo III-sensitive DNA damage in the livers of CPA-treated male rats; while Endo III-sensitive DNA damage was detected only in female rats treated with 25, 50, and 100 mg/kg/day CPA (A). All CPA doses resulted in significant increases in hOGG1-sensitive oxidative DNA damage in the livers of male rats; increases in hOGG1-sensitive DNA damage were detected only in female rats treated with 50 and 100 mg/kg/day CPA (B). *Significant at p≤0.05 relative to the vehicle control; error bars represent standard deviation. Please click here to view a larger version of this figure.
Tables 1&2. Histopathology analysis results. Since cytotoxicity can generate false-positive comet assay results, histopathological assessments for apoptotic and/or necrotic cells should be conducted for comet-positive tissues. In the current study, no CPA-induced hepatocyte apoptosis or necrosis was observed in livers of both male and female rats, which excluded the possibility of a false positive comet assay response.
| Lesion | Data | Dose CPA | ||||
| 0 mg/kg | 10 mg/kg | 25 mg/kg | 50 mg/kg | 100 mg/kg | ||
| Hepatocyte cytoplasmic vacuolization | Lesion Count | 1 | 1 | 5 | 5 | 6 |
| # Examined | 6 | 5 | 5 | 5 | 6 | |
| Lesion% | 17% | 20% | 100% | 100% | 100% | |
| Avg Severity | 1.0 | 1.0 | 1.2 | 1.8 | 2.0 | |
| Hepatocyte mitosis | Lesion Count | 0 | 0 | 5 | 5 | 6 |
| # Examined | 6 | 5 | 5 | 5 | 6 | |
| Lesion% | 0 | 0 | 100% | 100% | 100% | |
| Avg Severity | 0 | 0 | 1.0 | 1.4 | 1.8 | |
Table 1. Incidence of non-neoplastic lesions in livers of vehicle- and CPA-treated male F344 rats.
| Lesion | Data | Dose CPA | ||||
| 0 mg/kg | 10 mg/kg | 25 mg/kg | 50 mg/kg | 100 mg/kg | ||
| Hepatocyte cytoplasmic vacuolization | Lesion Count | 1 | 3 | 5 | 5 | 5 |
| # Examined | 5 | 5 | 5 | 5 | 5 | |
| Lesion% | 20% | 60% | 100% | 100% | 100% | |
| Avg Severity | 1.0 | 1.3 | 1.8 | 1.8 | 2.0 | |
| Hepatocyte mitosis | Lesion Count | 0 | 5 | 5 | 5 | 5 |
| # Examined | 5 | 5 | 5 | 5 | 5 | |
| Lesion% | 0 | 100% | 100% | 100% | 100% | |
| Avg Severity | 0 | 1.0 | 1.2 | 1.8 | 2.0 | |
| Hepatocyte Karyomegaly | Lesion Count | 0 | 0 | 5 | 5 | 5 |
| # Examined | 5 | 5 | 5 | 5 | 5 | |
| Lesion% | 0 | 0 | 100% | 100% | 100% | |
| Avg Severity | 0 | 1.0 | 1.2 | 1.8 | 2.0 | |
Table 2. Incidence of non-neoplastic lesions in livers of vehicle-and CPA-treated female F344 rats.
