Questo documento fornisce un test affidabile per segnalare il relativo espressione superficie cellulare di proteine di membrana, quali canali ionici espressi in cellule ricombinanti utilizzando la tecnologia di citometria a flusso esistente. I canali ionici sono proteine di membrana che formano pori che sono responsabili del controllo segnali elettrici dal gating il flusso di ioni attraverso la membrana cellulare. Essi sono classificati dal meccanismo di attivazione, la natura e la selettività di specie ioniche in transito attraverso il poro dove sono localizzate. A livello cellulare e dei tessuti, i flussi ionici macroscopici attraverso i canali ionici sono il prodotto di proprietà 1 biofisica (gating e permeazione), biochimici (fosforilazione), e biogenesi (sintesi, glicosilazione, il traffico, e la degradazione). Ciascuno di questi processi è unico per ogni tipo di canali ionici ed è ottimizzato per svolgere il ruolo fisiologico del canale ionico. Di conseguenza, alterazioni in nessuno di questi processi di precisione La attraverso unereditaria o una modificazione genetica, spesso definito come "canalopatia", può essere dannoso per l'omeostasi cellulare. È importante sottolineare che fornire l'importo "diritto" di canali ionici sulla superficie cellulare è fondamentale per l'omeostasi cellulare. Anche piccoli incrementi (guadagno-di-funzione) e lievi diminuzioni (perdita-di-funzione) in attività di canale ionico hanno il potenziale di causare una grave patologia nel corso della vita. Difetti nella consegna superficie cellulare dei canali ionici maturo è un importante determinante in numerosi canalopatie, come la fibrosi cistica (CFTR canale ionico) 2 e aritmie cardiache della forma lunga sindrome del QT (canali del potassio cardiaci) 3.
Canalopatie sono associati con cardiaca improvvisa morte 4. L'attuale diffusione a livello mondiale di tutti i canalopatie cardiache è pensato per essere di almeno 1: 2,000-1: 3.000 per individuo 5 e sono responsabili di circa la metà di improvvisa aritmica ca morte cardiacases 6. Disfunzione cardiaca voltaggio-dipendenti sodio, potassio, calcio-e canali ionici selettivi sono noti per svolgere un ruolo chiave in questo processo. Il 1.2 canali del calcio voltaggio-dipendenti L-tipo Ca V è tenuta ad avviare sincronizzato cuore la contrazione muscolare. Il cardiaca L-tipo Ca V 1.2 canali è un complesso proteico multi-subunità composto principale pore-forming Ca V α1 subunità e Ca V ß e Ca V α2δ1 subunità ausiliari 7-12. Si noti che la serie completa di subunità ausiliari è necessaria per produrre funzionali Ca V 1.2 canali a livello della membrana plasmatica e le interazioni dinamiche tra queste subunità sono essenziali per sostenere la funzione elettrica normale del cuore 13. Ca V ß promuove la superficie espressione cellulare di Ca V 1.2 canali attraverso un non-covalente nanomolari interazione idrofobica 14. Co-espressione del Ca V α2δ1 subunità wi° Ca V ß-bound V α1 Ca stimola l'espressione corrente di picco (da 5 a 10 volte) e promuove attivazione del canale a tensioni più negativi. Guadagno-di-funzione mutazioni della subunità pore-forming Ca V 1.2 sono stati associati con una forma di aritmia ventricolare chiamato la sindrome del QT lungo 15, mentre una serie di mutazioni puntiformi nelle tre principali subunità che formano il L-tipo Ca V 1.2 canali sono stati identificati nei soggetti affetti da aritmie del breve QT modulo sindrome di 16,17. I canali ionici sono proteine di membrana che possono essere studiati da un punto di vista biochimico (chimica delle proteine) o utilizzando gli strumenti di elettrofisiologia (macchine generatori di corrente) e spesso utilizzano questi approcci complementari. Elettrofisiologia, in particolare whole-cell patch-bloccaggio, è un approccio adatto per chiarire la funzione dei canali ionici 15, ma non è in grado di risolvere le modifiche nel traffico di proteine da cambiamenti nella loro biofisicaproprietà. Proteina chimica ha, tuttavia, spesso uso limitato a causa della relativamente bassa espressione di grandi proteine di membrana relativi alle proteine solubili piccoli. Robusti metodi high-throughput utilizzando la fluorescenza di lettura devono essere sviluppate al fine di affrontare specificamente i difetti di biogenesi di proteine che causano cambiamenti nell'espressione superficie cellulare dei canali ionici.
Citometria a flusso è una tecnologia impiegata in biofisica conteggio delle cellule, l'ordinamento, la rilevazione biomarker, e ingegneria proteica 18. Quando una soluzione campione di cellule vive o particelle viene iniettato in un citometro di flusso, le cellule sono ordinate in un unico flusso che può essere controllato di sistema di rilevamento della macchina (figura 1). Il primo citofluorimetro atto prodotto nel 1956 19 rilevato un solo parametro, ma moderni citofluorimetri avere più laser e rilevatori di fluorescenza che consentono il rilevamento di più di 30 parametri fluorescenti 20,21.Filtri e specchi (ottica di emissione) dirigono la diffusione luminosa o luce fluorescente di cellule ad una rete elettronica (fotodiodo e rilevatori) che convertono la luce in proporzione alla sua intensità. I dati digitali vengono analizzati utilizzando software specializzati e l'uscita principale viene visualizzato come un diagramma a punti 21.
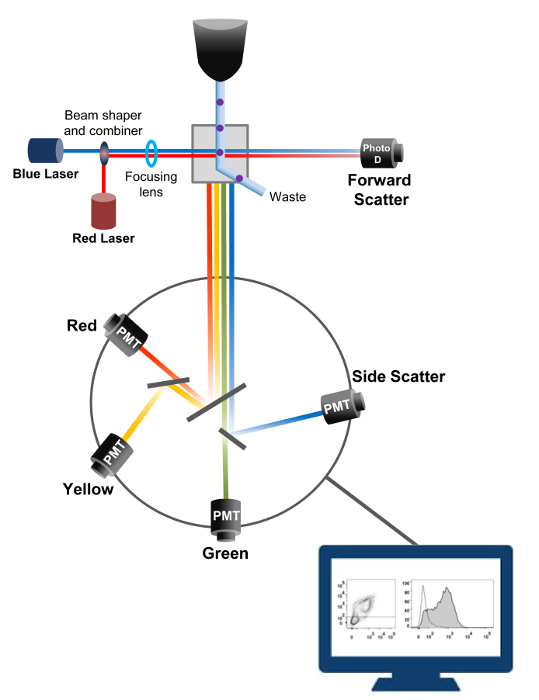
Figura 1:. Principi biofisici di citometria a flusso ordinamento celle singole sono spinti attraverso un ugello ad alta pressione all'interno di un flusso di liquido guaina che li muove attraverso uno o più punti di interrogatorio laser. Il fascio di luce viene deviato da cellule di passaggio e la luce raccolta in direzione avanti (Forward Scatter, FCS) viene inviata ad un fotodiodo che converte la luce in un segnale proporzionale alla dimensione della cella. La luce viene raccolta anche ad un angolo di 90 ° rispetto alla traiettoria laser e inviato ai rivelatori (chiamati anche fotomoltiplicatori (PMT)).Questa luce viene instradato attraverso specchi dicroici che permettono la rilevazione del segnale side scatter (SSC), che riflette la granularità all'interno delle cellule, e le emissioni fluorescenti se fluorocromi eccitati sono presenti nella cellula. Tre rivelatori (verde, giallo e rosso) sono rappresentati con diversi filtri passa-banda di lunghezza d'onda, permettendo la rilevazione simultanea di diversi fluorocromi. I diversi segnali vengono digitalizzati da un computer esterno e convertiti in dati che verranno analizzati per quantificare le caratteristiche delle cellule. Si prega di cliccare qui per vedere una versione più grande di questa figura.
La capacità high-throughput di citometri a flusso è stata sfruttata per quantificare l'espressione di membrana relativo di ricombinante wild-type e la tratta con deficit di voltaggio-dipendenti di tipo L Ca V 1.2 canali e subunità associati in cellule vive. cDNA costruisce coding per le proteine erano doppiamente etichettato avere simultaneamente un epitopo non fluorescente extracellulare che può essere rilevata da un anticorpo coniugato fluorescente impermeabile e un fluoroforo intracellulare che è costitutivamente fluorescente. Sia l'epitopo extracellulare, inserito in un loop extracellulare della proteina, e il fluoroforo intracellulare, inserito dopo il C-terminale, sono tradotti con la proteina. In questa serie di esperimenti, la proteina Ca V α2δ1 è stato progettato per esprimere un emoagglutinina extracellulare (HA) epitopo (YPYDVPDYA) rilevata da un impermeabile FITC (fluoresceina isotiocianato) coniugata anti-HA e mCherry come fluoroforo intracellulare intrinseca. Per determinare il relativo livello di espressione della superficie delle cellule del V α2δ1 mCherry-Ca proteina HA-tag, le cellule ricombinanti che esprimono la proteina di fusione sono state raccolte dopo la transfezione, e colorati con la FITC-coniugato topo monoclonale anti-HA tag epitopo gli anticorpiy (Figura 2). FITC è un composto fluorescente organico che è considerevolmente inferiore rispetto reporter enzimi e quindi improbabile interferire con la funzione biologica. mCherry- Ca V α2δ1-HA sovraespresso in Tsa-201cells, produce un significativo aumento di 3 log in fluorescenza FITC e mCherry fluorescenza appezzamenti bidimensionali 22. Dato che l'epitopo HA si trova nella porzione extracellulare della proteina, l'intensità di fluorescenza per FITC ottenuta in presenza di cellule intatte riflettono l'indice relativa della superficie cellulare espressione di proteine HA-tag. L'accessibilità del epitopo HA nei costrutti è sistematicamente convalidata misurando il segnale FITC dopo permeabilizzazione cellulare. Questa misura serve anche a corroborare l'espressione della proteina totale normalizzato dal momento che le intensità di fluorescenza relative per FITC stimati in cellule permeabilizzate sono qualitativamente paragonabili ai valori di fluorescenza relativi FOR mCherry misurato in condizioni permeabilizzate e non permeabilizzate 22,23. È importante notare che lo spettro di fluorescenza intrinseca è spostato verso valori più elevati dopo permeabilizzazione ma che l'unico valore di essere rilevato è la variazione di intensità di fluorescenza rispetto al costrutto controllo. variazioni relative l'intensità di fluorescenza per i costrutti di prova sono stimate utilizzando la Intensity ΔMean fluorescenza (ΔMFI) i valori per ciascun fluoroforo (mCherry o FITC). Gli esperimenti sono progettati per misurare l'intensità di fluorescenza della prova costrutto relativa all'intensità di fluorescenza del costrutto di controllo espressa nelle stesse condizioni sperimentali per limitare le variazioni nella fluorescenza intrinseca dell'anticorpo fluoroforo-coniugato. Due proteine di membrana sono stati studiati con successo utilizzando questo test: la subunità pore-forming del canale del calcio voltaggio-dipendenti di tipo L Ca V 1.2 14,22 e in una diversa serie diesperimenti, il extracellulare ausiliario Ca V α2δ1 subunità 22,23. Il seguente protocollo è stato utilizzato per determinare l'espressione superficie cellulare del Ca V α2δ1 subunità del cardiaca L-tipo Ca V 1.2 canale in condizioni di controllo e dopo mutazioni colpendo la modifica post-traslazionale del canale ionico. In condizioni sperimentali standardizzate, la fluorescenza superficie cellulare di FITC aumenta quasi linearmente con l'espressione di cDNA codificanti per le proteine α2δ1-HA mCherry-Ca V (Figura 5 dal riferimento 22).
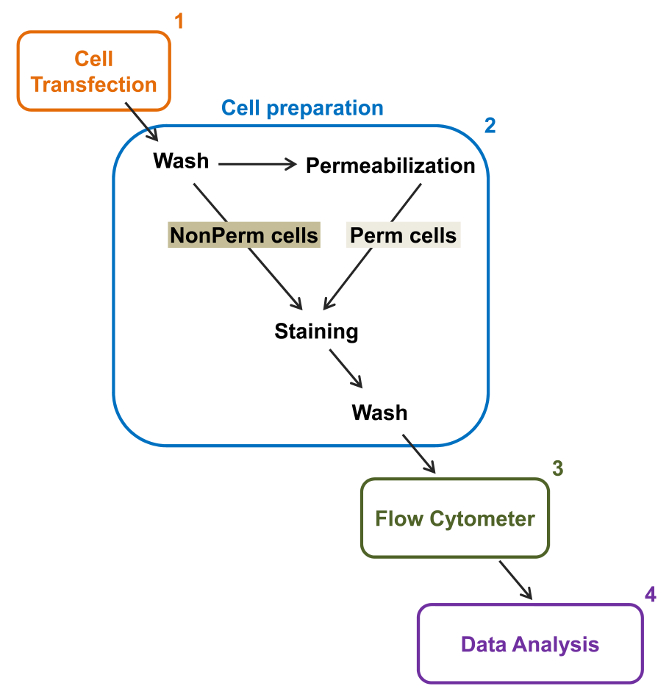
Figura 2:. Rappresentazione schematica di etichettatura totale e membrana in citometria a flusso protocollo sperimentale Lo schema illustra alcuni dei principali passi necessari per quantificare l'espressione relativa totale e superficie cellulare dei canali ionici ricombinanti di flOW citometria. Le cellule sono trasfettate con la costruzione doppia marcatura mCherry-Ca V α2δ1-HA in TSA-201 celle (1) e colorate prima o dopo permeabilizzazione (2). Dati multiparametrica vengono acquisiti in un citofluorimetro (3) per l'analisi multivariata (4). Si prega di cliccare qui per vedere una versione più grande di questa figura.
