I detta dokument ges en tillförlitlig analys för att rapportera den relativa cellytan uttryck av membranproteiner såsom jonkanaler som uttrycks i rekombinanta celler med användning av den befintliga flödescytometri teknik. Jonkanaler är porbildande membranproteiner som är ansvariga för styrning av elektriska signaler genom grind flödet av joner genom cellmembranet. De klassificeras av aktiveringsmekanismen, natur, och selektivitet jonslag transit genom poren där de är lokaliserade. Vid de cellulära och vävnadsnivåer, de makroskopiska jonflöden genom jonkanaler är en produkt av biofysiska (gating och permeation), biokemiska (fosforylering), och biogenes (syntes, glykosylering, trafficking, och nedbrytning) egenskaper 1. Var och en av dessa processer är unik för varje typ av jonkanaler och är optimerad för att uppfylla den fysiologiska rollen för jonkanalen. Följaktligen förändringar i någon av dessa finjusterade processer genom enärftlig eller genetisk modifiering, ofta kallad "channelopathy", kan vara skadliga för cell homeostas. Det är viktigt att betona att leverera "rätt" mängd av jonkanaler vid cellytan är kritisk för cell homeostas. Även små ökningar (vinst-of-funktion) och små minskningar (förlust-of-funktion) i jonkanalaktiviteten har potential att orsaka en allvarlig patologi under en livstid. Defekter i cellytan leverans av mogna jonkanaler är en viktig determinant i talrika channelopathy, såsom cystisk fibros (CFTR jonkanalen) 2 och hjärtarytmier av långt QT-syndrom form (hjärtkaliumkanaler) 3.
Channelopathy förknippas med hjärt plötslig död 4. Den nuvarande globala förekomsten av alla hjärt channelopathy tros vara åtminstone 1: 2,000-1: 3000 per individ 5 och svarar för ungefär hälften av plötslig arytmier hjärtdöd caSES 6. Dysfunktion i hjärtspänningskänsliga natrium-, kalium- och kalcium- selektiva jonkanaler är kända för att spela en nyckelroll i denna process. L-typ Ca V 1,2 spänningsstyrda kalciumkanal krävs för att initiera synkroniserad hjärtmuskeln sammandragning. Hjärt L-typ Ca V 1,2-kanal är en multi-subenhet proteinkomplex bestående av huvud porbildande Ca V α1 subenheten och Ca V ß och Ca V α2δ1 hjälp subenheter 7-12. Observera att full uppsättning av hjälp subenheter krävs för att producera funktionella Ca V 1.2 kanaler på plasmamembranet och dynamiska samspelet mellan dessa subenheter är nödvändiga för att stödja den normala elektriska funktionen av hjärtat 13. Ca V ß befrämjar cellytan uttryck av ca V 1.2 kanaler genom en icke-kovalent nanomolära hydrofob växelverkan 14. Samexpression av Ca V α2δ1 subenhet with Ca V ß bundna Ca V α1 stimulerar toppström uttryck (5-10-faldigt) och främjar kanalaktivering på mer negativa spänningar. Få-of-funktion mutationer av porbildande subenhet Ca V 1,2 har förknippats med en form av ventrikulära arytmier kallas långt QT-syndrom 15 medan en mängd punktmutationer i de tre huvudenheterna bildar L-typ Ca V 1,2 kanal har identifierats hos patienter som lider av arytmier av kort QT-syndrom blankett 16,17. Jonkanaler är membranproteiner som kan undersökas ur en biokemisk perspektiv (proteinkemi) eller med hjälp av elektrofysiologiska verktyg (strömgenererande maskiner) och ofta använder dessa kompletterande metoder. Elektrofysiologi, särskilt hel-cell patch-fastspänning, är en lämplig metod för att belysa funktionen av jonkanaler 15 men kan inte lösa modifikationer i proteinhandeln från förändringar i deras biofysiskaegenskaper. Proteinkemi har dock ofta begränsad användning på grund av den relativt låga uttrycket av stora membranproteiner i förhållande till mindre lösliga proteiner. Robusta hög genomströmning metoder med hjälp av fluorescens avläsning måste utvecklas för att specifikt behandla defekter i protein biogenes orsakar förändringar i cellytan uttryck av jonkanaler.
Flödescytometri är en biofysisk teknik som används i cellräkning, sortering, biomarkör upptäckt, och protein 18. När en provlösning av levande celler eller partiklar injiceras i en flödescytometer är cellerna ordnade till en enda ström som kan sonderas av maskinens detektionssystem (figur 1). Den första flödescytometer instrument som produceras i 1956 19 upptäckt bara en parameter men moderna flödescytometrar har flera lasrar och fluorescensdetektorer som medger detektering av mer än 30 fluorescerande parametrar 20,21.Filter och speglar (utsläpps optik) rikta ljusspridning eller fluorescerande ljus av celler till ett elektroniskt nätverk (fotodiod och detektorer) som omvandlar ljuset i förhållande till dess intensitet. Digitala data analyseras med användning av specialiserad mjukvara och primärutgången visas som ett punktdiagram 21.
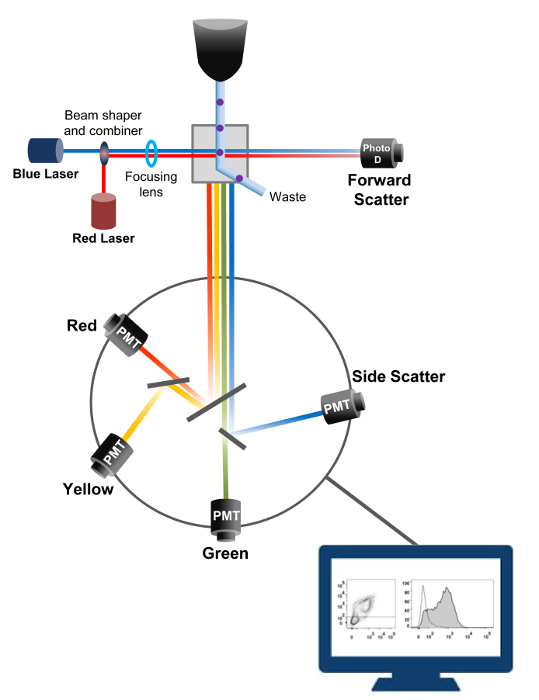
Figur 1:. Biophysical principerna för flödescytometri sortering Enstaka celler skjuts genom ett munstycke under högt tryck i en ström av omslutande vätska som förflyttar dem över en eller flera laserfrågepunkter. Ljusstrålen avlänkas av de passerande celler och ljuset uppsamlas i framåtriktningen (framåtspridning, FCS) sänds till en fotodiod som omvandlar ljuset till en signal som är proportionell mot storleken på cellen. Ljuset samlas också in vid en 90 ° vinkel i förhållande till laserbanan och sänds till detektorer (även kallade fotomultiplikatorer (PMT)).Detta ljus leds genom dikroiska speglar som tillåter detektering av sidospridningssignalen (SSC), vilket återspeglar den granularitet i cellerna, och de fluorescerande utsläppen om excite fluorokromer är närvarande i cellen. Tre detektorer (grön, gul och röd) är representerade med olika våglängd bandpassfilter, vilket gör att samtidig detektion av olika fluorokromer. De olika signalerna digitaliseras av en extern dator och omvandlas till data som kommer att analyseras för att kvantifiera egenskaper hos cellerna. Klicka här för att se en större version av denna siffra.
Hög kapacitet på flödescytometrar utnyttjades för att kvantifiera den relativa membran uttryck av rekombinant vildtyp och handel fattiga spänningskänsliga L-typ Ca V 1.2 kanaler och tillhörande subenheter i levande celler. cDNA konstruktioner coding för proteinerna dubbelt taggade att samtidigt ha en extracellulär icke-fluorescerande epitop som kan detekteras av en ogenomtränglig fluorescerande konjugerad antikropp och en intracellulär fluorofor som är konstitutivt fluorescerande. Både den extracellulära epitopen, insatt i en extracellulär slinga av proteinet, och den intracellulära fluorofor, införas efter C-terminalen, omräknas med proteinet. I denna serie av experiment, var Ca V α2δ1 protein konstruerad för att uttrycka ett extracellulärt hemagglutinin (HA) epitop (YPYDVPDYA) detekteras av en ogenomtränglig FITC (Fluoresceinisotiocyanat) -konjugerad anti-HA och mCherry som inneboende intracellulära fluoroforen. Att bestämma den relativa cellytan uttrycksnivån för mCherry-Ca V α2δ1 HA-märkt protein, rekombinanta celler som uttrycker fusionsproteinet skördas efter transfektion, och färgades med FITC-konjugerad monoklonal mus-anti-HA-epitop-taggen Antibody (Figur 2). FITC är en organisk fluorescerande förening som är betydligt mindre än enzym reportrar och därför inte så troligt att störa biologisk funktion. mCherry- Ca V α2δ1-HA överuttryckt i TSA-201cells, producerar en betydande 3-log ökning av FITC-fluorescens och mCherry fluorescens på tvådimensionella tomter 22. Med tanke på att HA-epitopen är belägen i den extracellulära delen av proteinet, fluorescensintensiteten för FITC erhölls i närvaro av intakta celler återspeglar den relativa index för cellytan uttryck av HA-märkt protein. Tillgängligheten till HA-epitopen i konstruktionerna systematiskt valideras genom mätning av FITC-signalen efter cell permeabilization. Denna åtgärd tjänar också till att bekräfta den normaliserade totala proteinuttryck eftersom de relativa fluorescensintensiteter för FITC beräknas i permeabiliserade celler är kvalitativt jämförbar med de relativa fluorescensvärdena for mCherry mätt under permeabiliserade och icke-permeabiliserade förhållanden 22,23. Det är viktigt att notera att den inneboende fluorescensspektrum skiftas mot högre värden efter permeabilisering men att det enda värde som rapporteras är den förändring i fluorescensintensitet jämfört med kontrollkonstruktionen. Relativa förändringar i fluorescensintensiteten för test konstruktioner uppskattas med hjälp av ΔMean Fluorescence Intensity (ΔMFI) värden för varje fluoroforen (mCherry eller FITC). Experiment har utformats för att mäta fluorescensintensiteten hos test konstruktionen i förhållande till fluorescensintensiteten hos kontroll konstruktet uttrycks under samma betingelser för att begränsa experimentella variationer i den inneboende fluorescensen av fluoroforen-konjugerade antikroppen. Två membranproteiner styrde studerades med användning av denna analys: det porbildande underenheten av L-typ spänningsstyrda kalciumkanal Ca V 1,2 14,22 och i en annan serie avexperiment, den extracellulära hjälp Ca V α2δ1 subenhet 22,23. Följande protokoll användes för att bestämma cellytan uttryck av Ca V α2δ1 subenheten av hjärt L-typ Ca V 1,2-kanal under kontrollbetingelser och efter mutationer som påverkar den posttranslationella modifieringen av jonkanalen. Under standardiserade experimentella betingelser, ökar cellytan fluorescens av FITC kvasi-linjärt med uttryck av cDNA som kodar för de mCherry-Ca V α2δ1-HA-proteiner (Figur 5 från referens 22).
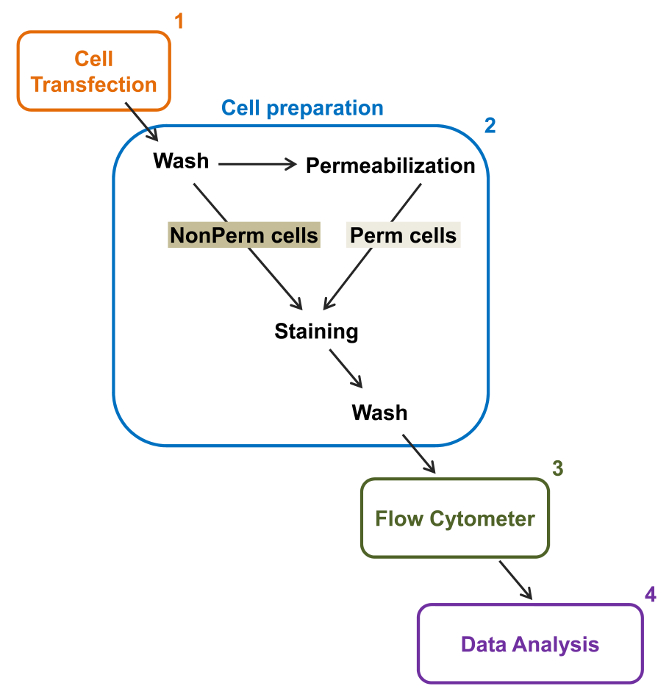
Figur 2:. Schematisk bild av den totala och membranmärkning i flödescytometri experimentella protokollet Systemet beskrivs några av de viktigaste stegen som krävs för att kvantifiera relativa totala och cellytan uttryck av rekombinanta jonkanaler från flow cytometry. Celler transfekteras med den dubbelmärkas konstruktion mCherry-Ca V α2δ1-HA i TSA-201-celler (1) och färgades före eller efter permeabilisering (2). FLERA uppgifter förvärvas i en flödescytometer (3) för multivariat analys (4). Klicka här för att se en större version av denna siffra.
