أوراق الذرة هي نموذج مثالي لدراسة تشكيل المجالات التنموية خلال التشكل، كما أن لديها حدود واضح بين النصل والغمد التي هي قابلة للتشريح الجيني (الشكل 1A). خلال المراحل الأولى من تطوير ورقة، عصابة خطية من الخلايا الصغيرة، والفرقة preligule (PLB)، الإضافي يقسم منشم ورقة إلى ما قبل النصل والغمد قبل المجالات. واللسيناء مثل هامشية والأذين الثلاثي تطوير من PLB (الشكل 1A، C، D). وقد حددت شاشات الوراثية الطفرات التي تعطل الحدود شفرة غمد. على سبيل المثال، liguleless1 المتنحية (LG1) الطفرات بحذف اللسيناء والأذين 1، 2، 3، 4 (الشكل 1B). في الموقع كشفت التهجين التي نص LG1 يتراكم في PLB واللسيناء الناشئة، مما يجعلها علامة ممتازة للتنمية اللسيناء 5، 6 (الشكل 1E).
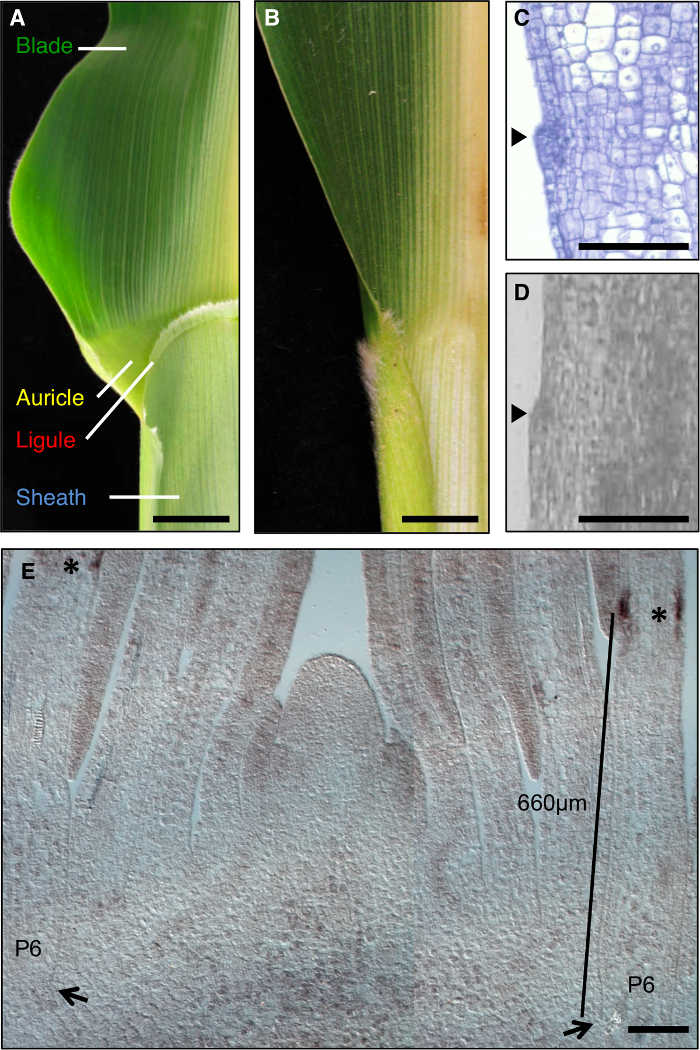
الشكل 1: من النوع البري وأوراق الذرة liguleless1-R. (أ) المنطقة بليد غمد حدود ناضجة ورقة من النوع البري تظهر الهياكل اللسيناء والأذن. (ب) المنطقة بليد غمد حدود ناضجة liguleless1-R ورقة تبين عدم وجود هياكل اللسيناء والأذن. قطعت الأوراق في A و B في نصف طول الضلع الأوسط. (C) الطولي القسم من خلال البرية من نوع ورقة منشم. وقد تم تجهيز عينة وملطخة للتحليل النسيجي. واللسيناء بدء واضح كما عثرة جاحظ من الطائرة من ورقة (رأس السهم). (D) طائفة طوليةايون من خلال البرية من نوع ورقة منشم. وقد تم تجهيز عينة لLM كما هو موضح في النص. يشير السهم الشروع اللسيناء. (E) LG1 في الموقع التهجين من تبادل لاطلاق النار مهيمنة المقطع الطولي الجانبي. وتشير العلامات النجمية تراكم نسخة LG1 في PLB من منشم ورقة P6. وتشير السهام قاعدة P6 منشم. يشير شريط القياس من قاعدة منشم إلى PLB. الحانات النطاق في A و B = 20 مم. الحانات النطاق في م = 100 ميكرون. تم تعديل هذا الرقم من الإشارة 6 (الجمعية الأمريكية حقوق الطبع والنشر علماء الأحياء النباتية). الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
في هذه الدراسة، كان يعمل LM RNA تسلسل لتحديد مجموعة من الجينات التي يتم التعبير عنها بشكل مختلف (DE) في شفرة غمد حدود نسبة إلى أجزاء أخرى من منشم ورقة وبيئة تطوير متكاملة الجينات ntify التي هي دي في المسوخ LG1-R قريبة من النوع البري الأشقاء. LM RNA تسلسل هو وسيلة لقياس تراكم نص في خلايا معينة أو المجالات الخلوية 7. نظم LM الجمع بين الليزر والمجهر مع الكاميرا الرقمية. هي التي شنت الأنسجة مقطوع على الشرائح وعرضها من خلال المجهر. البرنامج LM عادة ما تضم أدوات الرسم التي تسمح للمستخدم لمخطط أي منطقة تم اختيارها لتسليخ مجهري. تخفيضات ليزر على طول الخط، والأنسجة المختارة قفزت خارج الشريحة وفي أنبوب معلق فوق الشريحة. LM يسمح للمستخدم لmicrodissect مجالات محددة، بما في ذلك طبقات معينة من الخلايا وحتى الخلايا وحيدة 8 و 9. RNA ومن ثم يمكن استخراجها من أنسجة microdissected. وفي وقت لاحق، المكون RNA يليها يستخدم الجيل القادم التسلسل تسلسل مكتبات كدنا] المتولدة من الحمض النووي الريبي المستخرج 10،= "XREF"> 11.
المزايا الرئيسية لLM RNA وما يليها هي القدرة على قياس تراكم نسخة في مجالات محددة بدقة والقدرة على محة عن Transcriptome على كامل في وقت واحد (7). هذه التقنية هي مناسبة بشكل خاص لسبر الأحداث التنموية في وقت مبكر حيث المنطقة من اهتمام في كثير من الأحيان المجهرية. وقد استخدمت الدراسات السابقة LM جنبا إلى جنب مع تكنولوجيا متطورة لدراسة العمليات التنموية في محطات 9 و 12 و 13. RNA تسلسل لديه ميزة قياس النصوص عبر مجموعة ديناميكية واسعة، بما في ذلك الجينات منخفضة أعرب، وليس مطلوبا من المعلومات تسلسل مسبق 10 و 11. وعلاوة على ذلك، LM RNA تسلسل لديه القدرة على تسليط الضوء على جينات هامة تنمويا قد غاب في شاشات الطفرات بسبب التكرار الجيني أو إلى الفتك من خسارة من-وظيفة متحولة.
جينات هامة تنمويا، مثل sheath1 ضيقة (NS1) وعلى شكل كوب cotyledon2 (cuc2)، وغالبا ما يكون أنماط التعبير محددة من واحد أو عدد قليل من الخلايا 17، 18، 19، 20. وأعرب كثيرون فقط خلال مراحل النمو المبكرة وليس في الجهاز ناضجة. عندما يتم تحليل أجهزة كاملة أو المجالات كبيرة، وتضعف هذه النصوص خلية محددة، ولا يجوز الكشف في التحليلات التقليدية أكثر. من خلال السماح تحليلات مجالات محددة بدقة، وتمكن LM RNA تسلسل هذه الجينات الأنسجة محددة يتم تحديدها وكميا.
ومن العوامل الحاسمة في نجاح هذه التجارب هو موضح هنا تحليل النسيجي الدقيق التي وجهت اختيار المرحلة التنموية المناسبة والمجال لتحليل، وmeasureme دقيقةالإقليم الشمالي من المجالات خلية الأنسجة لLM. لضمان المجالات تعادل وأخذت عينات لجميع مكررات، تم جمع الأنسجة من براعم نبات في هذه المرحلة التنموية نفسها، وكان قياس المجالات microdissected النسبية إلى المعالم المورفولوجية مثل اللسيناء الناشئة (الشكل 2). ومن المعروف أن يتم التعبير عن بعض الجينات في التدرج من طرف إلى قاعدة ورقة. عن طريق قياس المجالات دقيقة، والتباين بسبب أخذ العينات من مواقع مختلفة على طول المحور الداني القاصي ورقة كان في حده الأدنى (الشكل 3A). بواسطة microdissecting المجالات من نفس الحجم، التخفيف من النصوص خلية محددة تم تخفيض أيضا (الشكل 3B) الاختلاف يرجع إلى التفاضلية. واستخدمت المقاطع الطولية الجانبية للقمة تبادل لاطلاق النار لجميع microdissections. هذه هي المقاطع التي هي عمودي على محور الضلع الأوسط الهامش (الشكل 4). باستخدام أقسام الوحيدة التي تشمل SAM يضمن أن المناطق الجانبية ما يعادلويتم تحليل براعم نبات.
في عينات المصنعة ومقطوع عن LM، أول علامة الصرفي من ثمرة اللسيناء هو نتوء على الجانب متجه نحو المحور بسبب انقسامات الخلية periclinal في البشرة متجه نحو المحور (1D الشكل، الشكل 2). تقرر أن اللسيناء الناشئة يمكن تحديدها بشكل موثوق به في فترة تتابع النمو 7 براعم نبات المرحلة. كنا مهتمين الجينات وأعرب في المنطقة اللسيناء بأكملها، بما في ذلك اللسيناء الناشئة والخلايا البعيدة على الفور من شأنها أن تشكل الأذن. من أجل ضمان أن التحديدات الأنسجة تعادل بذلت، تم استخدام عثرة اللسيناء كمعلم الصرفي واختير مستطيل 100 ميكرون تركزت على نتوء اللسيناء لLM (الشكل 2A، 2B). وقد تم اختيار مستطيلات يعادل الحجم من قبل شفرة وقبل غمد من نفس براعم نبات.
قدمت تحليلات للنباتات معدلة وراثيا liguleless وشال مختلفةنجى. LG1-R المسوخ لا تشكل اللسيناء، لذلك لا يمكن استخدام هذه الميزة المورفولوجية لتحديد المنطقة لLM. بدلا من ذلك، تم تحديد مجال تراكم نسخة LG1 في البرية من نوع نبات براعم، وأنها حددت المنطقة التي من شأنها أن تشمل هذا المجال. تم إجراء هذه التحاليل الأولية على الشتلات من نفس زراعة مثل استخدمت في التحليل النهائي، لأن العمل السابقة قد أظهرت أن موقع PLB يختلف باختلاف ظروف النمو. في الموقع أشار التهجين أن النصوص LG1 تتراكم في PLB من P6 رقة براعم (الشكل 1E). اخترنا مجال 400-900 ميكرون من قاعدة براعم نبات أن يشمل مجال التعبير LG1 (المستطيلات الأرجواني، الشكل 2A) واستولت على هذه المناطق ما يعادلها من النوع البري والنباتات LG1-R. لتقليل التباين في ظروف الخلفية والنمو الوراثية عند مقارنة transcripواستخدمت تراكم تي في LG1-R والنباتات البرية من نوع، فصل العائلات من المسوخ والبرية من نوع الأشقاء.
