Genetik diseksiyon (Şekil 1A) için uygun olan bıçak ve kılıf arasında belirgin bir sınır olduğu gibi mısır yaprağı, morfolojilerinden sırasında gelişimsel alanların oluşumunu incelemek için ideal bir model. yaprak gelişimi, küçük hücrelerden oluşan bir lineer grubun ilk aşamalarında, preligule bandı (PLB), pre-bıçak ve ön kılıf etki alanlarına yaprak primordium'u subdivides. Bir saçak gibi ligule ve üçgen kulakcık PLB (Şekil 1A, C, D) gelişir. Genetik ekranlar bıçak kılıf sınırını bozan mutasyonlar saptanmıştır. Örneğin, resesif liguleless1 (LG1) mutasyonları ligule kulakçıkların, 1, 2, 3, 4 (Şekil 1B) silin. İn situ hibridizasyon LG1 transkript P birikir ortayaLB ve ligule gelişme 5, 6 (Şekil 1E) için o mükemmel bir işaretleyici yaparak, ligule ortaya çıkıyor.
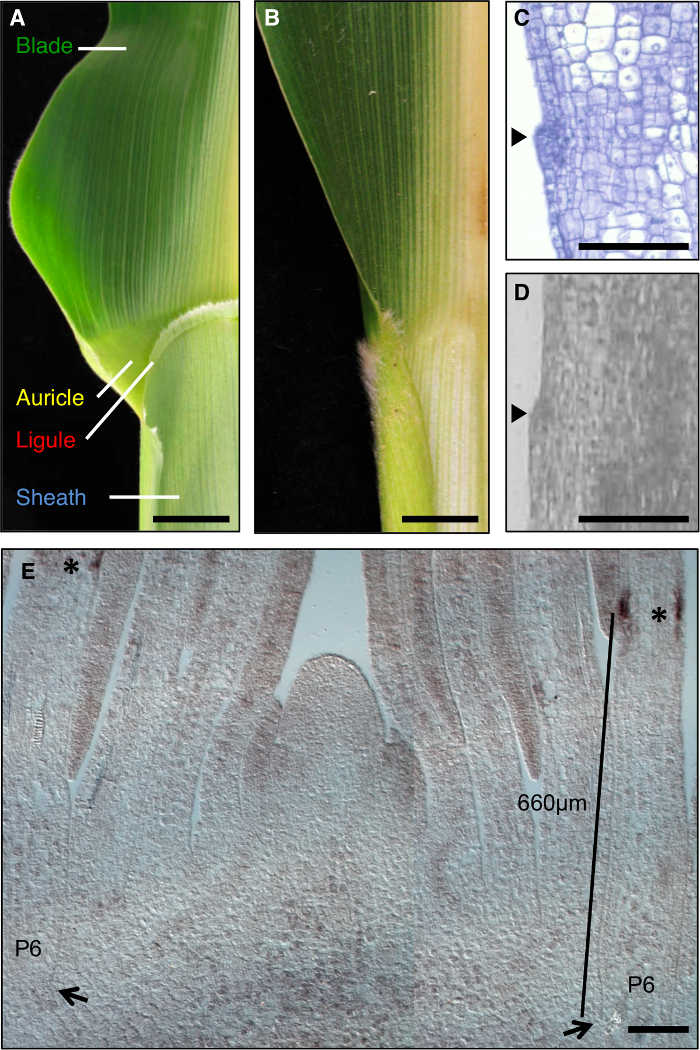
Şekil 1: vahşi tip ve liguleless1-R mısır yaprakları. Ligule ve kulak kepçesi yapıları gösteren, olgun, vahşi tip yaprak (A) Bıçak kaplama sınır bölgesi. (B) ligule ve kulak kepçesi yapıların olgun liguleless1-R yaprak gösteren yokluğunda Bıçak kılıf sınır bölgesi. A ve B Yapraklar yaprak orta damarı boyunca yarıya edilmiştir. Vahşi tip yaprak primordiumunun yoluyla (C) Boyuna kesit. Örnek işleme ve histolojik analiz için boyanmıştır edilmiştir. başlatılması ligule yaprak (ok başı) düzleminden dışarı çıkıntı yapan bir tümsek belirgindir. (D) Boyuna mezhepvahşi tip yaprak primordiumunun yoluyla iyon. metinde tarif edildiği gibi Örnek LM için işlenmiştir. Ok başı ligule başlatılması gösterir. Sürgün ucuna uzunlamasına yanal bölümün in situ hibridizasyon (E) LG1. Yıldız P6 yaprak primordiumunun PLB de LG1 transkript birikimi gösterir. Oklar P6 primordium'un tabanını gösterir. Bar PLB için primordium'un tabanından ölçümünü gösterir. A ve B Ölçek çubukları 20 mm =. CE Ölçek çubukları = 100 mikron. Bu rakam referans 6 (Bitki Biyologlar Telif Amerikan Derneği) modifiye edilmiştir. Bu rakamın büyük halini görmek için lütfen buraya tıklayınız.
Bu çalışmada, LM RNA Seq yaprak primordiumunun ve ide diğer bölgelerine bıçak-kılıf sınır nispetle farklı ifade edilen genlerin bir paketi (DE) tanımlamak için kullanılmıştır vahşi tip kardeşleri göre LG1-R mutantlar DE vardır ntify genler. LM RNA Seq özel hücreler ya da hücre alan 7 transkript birikimi ölçülmesi için bir yöntem olup. LM sistemleri lazer ve bir dijital kamera ile bir mikroskop birleştirir. Kesitli doku slaytlar üzerine monte edilir ve mikroskop aracılığıyla görüntülenebilir. LM yazılım genellikle kullanıcı mikrodiseksiyon seçilen herhangi bir bölgeyi anahat izin çizim araçları içerir. hat boyunca lazer kesim ve seçilen doku slayt kapalı mancınık ve slayt üzerinde asılı bir tüp içine almaktadır. LM kullanıcıya özel hücre katmanları ve hatta tek hücreler 8, 9 da dahil olmak üzere hassas etki, microdissect sağlar. RNA daha sonra microdissected dokusundan elde edilebilir. Daha sonra, RNA-Seq bileşen çıkarılan RNA 10 üretilen cDNA kütüphaneleri sıralamak için yeni nesil dizileme kullanır,= "xref"> 11.
LM RNA seq Anahtar avantajları kesin olarak tanımlanmış alanlarda transkript birikimi ölçmek yeteneği ve tüm transkriptom aynı anda 7 profile kapasite vardır. teknik faiz bölge genellikle mikroskobik erken gelişim olayları sondalama için uygundur. Daha önceki çalışmalar, LM bitkiler 9, 12, 13, gelişim süreçleri incelemek için mikrodizi teknolojisi ile birlikte kullanmıştır. RNA Seq düşük ifade edilen genler de dahil olmak üzere, geniş bir dinamik aralık, arasında transkript miktarının avantajı vardır ve önceki dizi bilgisi, 11 10 gerekli değildir. Ayrıca, LM RNA Seq nedeniyle-of-zarara genetik fazlalık veya öldürücülüğü için mutagenez ekranlarında gözden kaçabilir gelişimsel olarak önemli genlerin vurgulamak için potansiyele sahiptirfonksiyon mutant.
Bu tür dar bir sheath1 (NS1) ve kupa şekilli cotyledon2 (CUC2) halinde gelişimsel olarak önemli genler, genellikle sadece bir spesifik ekspresyon modelleri ya da birkaç hücre 17, 18, 19, 20 sahiptir. Çoğu sadece erken gelişim evrelerinde ve olgun bir organ olarak ifade edilir. Bütün organlar ya da büyük etki incelendiğinde, bu hücreye spesifik transkriptler seyreltilmiş ve daha geleneksel analizlerde tespit edilemez. kesin olarak tanımlanmış etki analiz edilmesine izin vererek, LM RNA Seq bu dokuya özgü genler belirlendi ve miktarları tayin edilmesini sağlar.
Burada tarif edilen deneylerde başarısında önemli faktörlerdir bir analiz için uygun gelişim aşamasında ve etki alanının seçimi güdümlü tam histolojik analiz ve hassas measureme vardıLM hücre dokusunu etki nt. Eş etki her kopya için örneklendi emin olmak için, doku, aynı gelişim aşamasında, yaprak primordia toplandı ve microdissected etki, ortaya çıkan ligule (Şekil 2) gibi morfolojik noktalara göre ölçülmüştür. Bazı genler, yaprak tabanına ucundan gradyanı olarak ifade edilmiştir bilinmektedir. Nedeniyle yaprak proksimal-distal ekseni boyunca farklı yerlerden numune hassas etki, varyasyon ölçerek en az (Şekil 3A) tutuldu. Aynı boyutta etki microdissecting ile nedeniyle değişkenlik hücreye özel transkriptlerin seyreltme (Şekil 3B) indirgenerek diferansiyel. Atış apeksinin yan uzunlamasına kesitleri, tüm microdissections kullanılmıştır. Bunlar (Şekil 4) yaprak orta damarı-margin eksenine dik bölümler vardır. SAM içeren tek bölümleri kullanarak o eşdeğer yanal bölgeleri sağlaryaprak taslakları analiz edilir.
İşlenmiş ve LM için kesitli numunelerde, ligule akıbet ilk morfolojik işareti nedeniyle adaxial epidermis (Şekil 1D, Şekil 2) periclinal hücre bölünmeleri adaxial tarafında bir darbedir. Gelişmekte olan ligule güvenilir plastochron 7 aşamalı yaprak primordiasında tespit edilebilir o belirlenmiştir. Biz gelişmekte olan ligule ve kulak kepçesi oluşturacaktır hemen uzak hücreleri dahil olmak üzere tüm ligule bölgede ifade genler, ilgilenmişlerdir. Eşdeğer doku seçimi yapılırken sağlamak amacıyla, ligule yumru morfolojik işareti olarak kullanıldı ve ligule yumru merkezli 100 um dikdörtgen LM (Şekil 2A, 2B) için seçildi. Ön bıçak ve ön kılıfın aynı büyüklükte dikdörtgenler aynı yaprak primordia seçilmiştir.
liguleless mutant bitkilerin analizi Farklı challe sunulmaktadırnge; LG1-R mutantlar bu nedenle bu morfolojik özellik LM için bölgeyi seçmek için kullanılan olamazdı bir ligule, oluşturmazlar. Bunun yerine, vahşi tip yaprak primordia olarak LG1 transkript birikimi alanı tespit edildi ve bu da kapsamak olan bir bölge tanımlanmıştır. son tahlilde için kullanılan önceki çalışma PLB yeri büyüme koşullarına bağlı olarak değişir göstermiştir beri bu ön analizler, aynı dikimden itibaren fidelerin üzerinde gerçekleştirilmiştir. In situ hibridizasyon LG1 transkript PLB P6 yaprak primordia (Şekil 1 E) birikir olduğunu göstermiştir. Biz LG1 ifadesinin alanını (mor dikdörtgenler, Şekil 2A) kapsıyor ve yabani tip ve LG1-R bitkilerden bu eşdeğer bölgeleri ele yaprak primordiasında tabanından bir alanı 400-900 mikron seçilmiş. transkripsiyonu karşılaştırırken genetik altyapı ve büyüme şartlarına varyasyonu en aza indirmek içinLG1-R ve vahşi tip bitkiler, mutant ve vahşi tipli kardeşlerinden ailelerini ayırma t birikimi kullanılmıştır.
