Bladet majs är en idealisk modell för att studera bildandet av utvecklingsområden under morfogenes, eftersom den har en tydlig gräns mellan bladet och manteln som är mottaglig för genetisk dissektion (Figur 1A). Under de tidiga stadierna av bladutveckling, en linjär band av mindre celler, den preligule bandet (PLB), delar upp blad primordium in i pre-bladiga och pre-slida domäner. En frans-liknande ligule och triangulära flikar utvecklas från PLB (Figur 1A, C, D). Genetiska skärmar har identifierat mutationer som stör blad manteln gränsen. Till exempel, recessiv liguleless1 (LG1) mutationer radera ligule och förmaksöronen 1, 2, 3, 4 (Figur 1B). In situ hybridisering visade att LG1 avskrift ackumuleras i PLB och nya ligule, vilket gör den till en utmärkt markör för ligule utveckling 5, 6 (figur 1E).
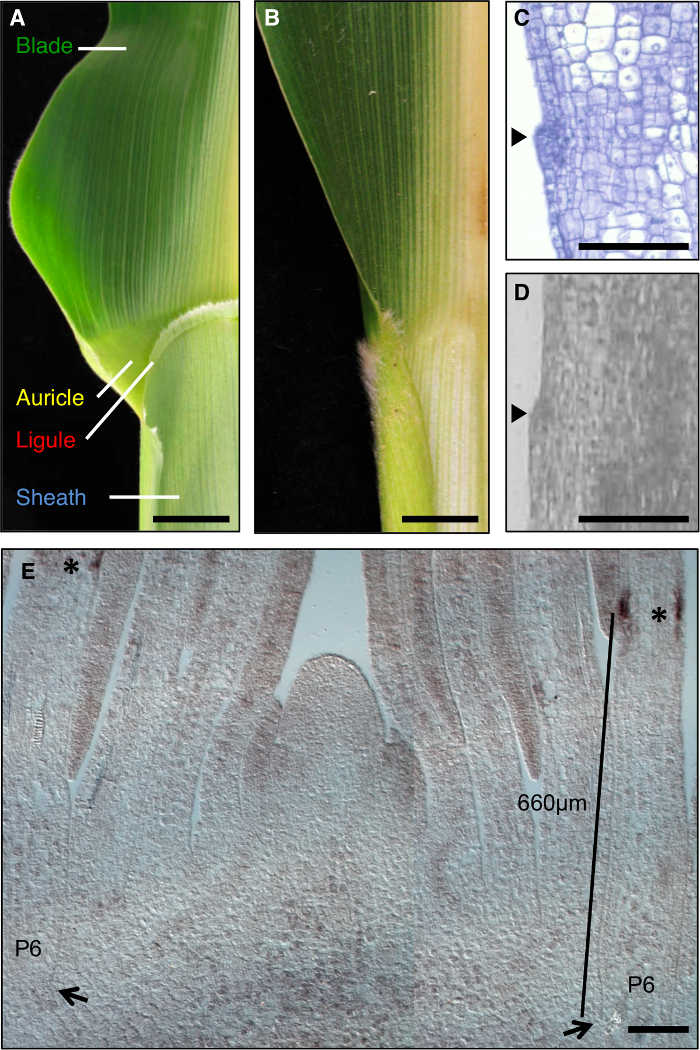
Figur 1: Vildtyp och liguleless1-R majsblad. (A) Blade-mantel gränsområdet av mogen vildtyp blad visar ligule och ytteröra strukturer. (B) Blade-mantel gränsområdet av mogen liguleless1-R blad som visar frånvaro av ligule och ytteröra strukturer. Bladen i A och B har halverats längs Mittnerven. (C) Längdsnitt genom vildtyp leaf primordium. Prov har behandlats och färgades för histologisk analys. Den initierande ligule framgår som en bula som skjuter ut från planet för blad (pilspets). (D) Longitudinell sektjon genom vildtyp leaf primordium. Provet har bearbetats för LM som beskrivs i texten. Pilspets indikerar initiering ligule. (E) LG1 in situ hybridisering av skott apex sidolängdsnitt. Asterisker indikerar LG1 avskrift ansamlingen på PLB av P6 blad primordium. Pilar anger basen av P6 primordium. Bar indikerar mätning från basen av primordium till PLB. Skala barer i A och B = 20 mm. Skala barer i CE = 100 pm. Denna siffra har modifierats referens 6 (Copyright American Society of Plant Biologists). Klicka här för att se en större version av denna siffra.
I denna studie var LM RNA-Seq användas för att identifiera en uppsättning gener som är differentiellt uttryckta (DE) vid bladets-mantelgräns i förhållande till andra delar av blad primordium och till IDE ntify gener som är DE LG1-R-mutanter i förhållande till vildtyp syskon. LM RNA-Seq är en metod för att kvantifiera avskrift ackumulering i specifika celler eller cellulära domäner 7. LM system kombinerar en laser och ett mikroskop med en digital kamera. Sektionerade vävnaden monteras på objektglas och betraktas genom mikroskopet. LM Programvaran innehåller typiskt ritverktyg som gör det möjligt för användaren att beskriva någon utvald region för microdissection. Laser skär längs linjen, och den valda vävnaden slungades från glaset och in i ett rör upphängd ovanför bilden. LM tillåter användaren att microdissect exakta domäner, inklusive specifika cellskikt och även enstaka celler 8, 9. RNA kan sedan extraheras från microdissected vävnaden. Därefter använder RNA-Seq komponent nästa generations sekvensering för att sekvensera cDNA-bibliotek som genereras från det extraherade RNA 10,= "xref"> 11.
Viktigaste fördelarna med LM RNA-punkter är förmågan att kvantifiera avskrift ackumulering i exakt definierade domäner och förmågan att profilera hela transkriptom samtidigt 7. Tekniken är särskilt lämpad för att sondera tidiga utvecklings evenemang där regionen av intresse är ofta mikroskopiska. Tidigare studier har använt LM kombinerat med microarray-teknik för att studera utvecklingsprocesser i växter 9, 12, 13. RNA-Seq har fördelen att kvantifiera transkript över ett brett dynamiskt omfång, inklusive lågt uttryckt gener och före sekvensinformation krävs inte 10, 11. Dessutom har LM RNA-Seq potential att lyfta fram utvecklings viktiga gener som kan missas i mutagenes skärmar på grund av genetiska redundans eller för dödlighet av förlusten-of-funktion mutanten.
Utvecklings viktiga gener, såsom smala sheath1 (NS1) och skålformad cotyledon2 (CUC2), har ofta specifika uttrycksmönster för bara en eller ett fåtal celler 17, 18, 19, 20. Många uttrycks endast under tidiga utvecklingsstadier och inte i den mogna organet. När hela organ eller stora domäner analyseras, är dessa cellspecifika transkript utspädd och kan inte upptäckas i mer konventionella analyser. Genom att tillåta analyser av exakt definierade domäner, LM RNA-Seq möjliggör dessa vävnadsspecifika gener kan identifieras och kvantifieras.
Avgörande faktorer för framgång de experiment som beskrivs här var en grundlig histologisk analys som styrt valet av lämplig utvecklingsstadiet och domän för analys, och exakt MeasureMent av cellvävnads domäner för LM. För att säkerställa att motsvarande domäner samplades för alla replikat, ades vävnad uppsamlades från blad primordia vid samma utvecklingsstadium och de microdissected domänerna mättes i förhållande till morfologiska landmärken, såsom den framväxande ligule (Figur 2). Det är känt att vissa gener uttrycks i en gradient från spetsen till basen av bladet. Genom att mäta exakta domäner, variation på grund av provtagning från olika platser längs blad proximal-distal axel hölls till ett minimum (figur 3A). Genom microdissecting domäner av samma storlek, för att variation på grund differentiell utspädning av cellspecifika transkript minskade också (figur 3B). Laterala längsgående sektioner av shoot apex användes för alla microdissections. Dessa är delar som är vinkelräta mot Mittnerven marginal axeln (Figur 4). Genom att bara använda delar som inkluderar SAM säkerställer att motsvarande sidoområdena avblad primordia analyseras.
I prover som behandlas och sektione för LM, är den första morfologiska tecken på ligule utväxt en bula på adaxial sidan på grund av periclinal celldelningar i adaxial epidermis (Figur 1D, Figur 2). Det konstaterades att den framväxande ligule kan tillförlitligt identifieras på plastochron 7 skede blad primordia. Vi var intresserade av gener som uttrycks i hela ligule regionen, inklusive den framväxande ligule och cellerna omedelbart distala som ska ligga ytterörat. I syfte att säkerställa att motsvarande vävnads urval gjordes, var ligule bula som används som en morfologisk landmärke och en 100 | j, m rektangel centrerad på ligule bump valdes för LM (figur 2A, 2B). Ekvivalenta stora rektanglar av pre-bladet och pre-mantel valdes från samma blad primordia.
Analyser av liguleless muterade växter presenterade en annan ChalleNBE; LG1-R-mutanter inte bildar en ligule, därför denna morfologiska funktionen inte kan användas för att välja region för LM. I stället var domänen för LG1 avskrift ackumulering i vild-typ blad primordia bestämdes och en region som skulle omfatta detta område definierades. Dessa preliminära analyser utfördes på plantor från samma plantering som användes för den slutliga analysen, eftersom tidigare arbete har visat att placeringen av PLB varierar beroende på odlingsbetingelser. In situ hybridisering indikerade att LG1 transkript ackumuleras i PLB av P6 blad primordia (figur 1E). Vi valde en domän 400-900 pm från basen av bladet primordia som omfattade domän LG1 uttryck (lila rektanglar, figur 2A) och fångade dessa motsvarande regioner från vild-typ och LG1-R växter. För att minimera variationer i genetisk bakgrund och tillväxtbetingelser när man jämför transcript ackumulering i LG1-R och vildtypsväxter, segregerande familjer mutanter och vildtyp syskon användes.
