Wechselwirkung zwischen Molekülen ist die Basis der Natur. Daher Wissenschaftler in vielen Bereichen der Grundlagen- und angewandten Forschung versuchen die grundlegenden Prinzipien der molekularen Wechselwirkungen verschiedener Arten zu verstehen. Micro Thermophorese (MST) können Wissenschaftler die schnelle, präzise und kosteneffiziente und qualitätskontrollierte Charakterisierung von molekularen Wechselwirkungen in Lösung durchzuführen, mit einer freien Wahl von Puffern. Es gibt bereits mehr als 1.000 Publikationen MST mit, ab 2016 allein, beschreibt verschiedene Arten von Analysen, einschließlich Bibliothek Vorführungen, Ereignis Validierungen Bindung, kompetitive Assays, und Experimente mit mehreren Bindungspartnern 1-8. Im Allgemeinen erlaubt MST die Untersuchung der klassischen Bindungsparameter wie Bindungsaffinität (pM bis Mm), Stöchiometrie und Thermodynamik, der jede Art von molekularen Wechselwirkungen. Ein großer Vorteil der MST ist die Fähigkeit, Bindungsereignisse unabhängig von der Größe der Interaktionspartner zu studieren. Auch Heraus-fordernd Wechselwirkungen zwischen kleinen Nukleinsäure-Aptamere (15-30 nt) und Ziele wie kleine Moleküle, Medikamente, Antibiotika oder Metaboliten quantifiziert werden können.
Aktuelle state-of-the-art Technologien Aptamer-Target – Wechselwirkungen sind entweder im Labor intensiv und sehr komplex zu charakterisieren oder scheitern Aptamer-kleines Molekül 9,10 Wechselwirkungen zu quantifizieren. Oberflächenplasmonresonanz (SPR) -basierte Assays 11,12 und wirklich markierungsfreie kalorimetrischen Ansätze wie Isothermen Titrations – Kalorimetrie (ITC) 13-15, isokratische Elution 16, Gleichgewicht Infiltrations 17,18, in-line Sondieren 19, Gel- Shift – Assays, Stopped fl ow Fluoreszenz- Spektroskopie 20,21, Fluoreszenzanisotropiemessungen (FA) 22,23, Einzelmolekül – Fluoreszenz – Bildgebung 24,25 und Bio-Schicht – Interferometrie (BLI) 26 sind ebenfalls entweder ungenau oder nicht kompatibel mit Aptamer-kleines Molekül Wechselwirkungen. Andere principal Probleme dieser Verfahren sind eine geringe Empfindlichkeit, hohe Probenverbrauch, Immobilisation, Massentransportbeschränkungen auf Oberflächen und / oder Puffer Einschränkungen. Nur wenige dieser Technologien bieten integrierte Kontrollen für die Aggregation und Adsorption Effekte.
MST stellt ein leistungsfähiges Werkzeug für Wissenschaftler , diese Einschränkung zu überwinden , um die Wechselwirkungen zwischen Aptamere und kleine Moleküle , 27-29, sowie andere Ziele, wie Proteine 30-33 zu studieren. Die Technologie beruht auf der Bewegung von Molekülen durch Temperaturgradienten. Diese gerichtete Bewegung, die so genannte "Thermophorese," hängt von der Größe, der Ladung und Hydrathülle des Moleküls 34,35. Die Bindung eines Liganden an das Molekül direkt an mindestens einen dieser Parameter zu verändern, in einer veränderten thermophoretische Mobilität führt. Liganden mit kleinen Größen haben nicht erhebliche Auswirkungen in Bezug auf die Größenänderung von der ungebundenen zum gebundenen Zustand, aber sie können dr haben amatic Auswirkungen auf die Hydrathülle und / oder Ladung. Die Veränderungen in der thermophoretische Bewegung der Moleküle nach der Wechselwirkung mit dem Bindungspartner ermöglicht die Quantifizierung der Grundbindungsparameter 2,7,34,36,37.
Wie in 1A dargestellt ist , besteht der MST Vorrichtung eines Infrarotlasers auf die Probe innerhalb der Glaskapillaren fokussiert die gleiche Optik wie für die Fluoreszenzdetektion. Die thermophoretischer Bewegung von Proteinen über die Eigenfluoreszenz von Tryptophane 6 oder eines fluoreszierend 3,8 Interaktionspartner markiert überwacht werden kann , während der Laser einen Temperaturgradienten (AT von 2-6 ° C) herstellt. Die sich ergebende Temperaturdifferenz im Raum, & Delta; T, führt zur Verarmung oder Anreicherung von Molekülen im Bereich der erhöhten Temperatur, die durch die Soret quantifiziert werden kann Koeffizienten (S T):
g "/>
c hot stellt die Konzentration in dem erwärmten Bereich, und c kalt ist die Konzentration in dem anfänglichen Kaltbereich.
Wie in 1B gezeigt , um eine typische MST Versuchsergebnisse in einer MST Bewegungsprofil (Zeitverlauf), die aus verschiedenen Phasen, die durch ihre jeweiligen Zeitrahmen getrennt werden können. Die anfängliche Fluoreszenz wird in den ersten 5 s in Abwesenheit des Temperaturgradienten gemessen, um die genaue Start Fluoreszenz zu definieren und für Photobleaching oder photoenhancement zu überprüfen. Der Temperatursprung (T-Jump) repräsentiert die Phase, in der die Fluoreszenzänderungen vor thermophoretischer Bewegung. Diese anfängliche Abnahme der Fluoreszenz hängt von wärmeabhängige Veränderungen von fl uorophore Quantenausbeute. Die Thermophorese Phase folgt, in der die Fluoreszenz abnimmt (oder zunimmt) aufgrund der thermophoretische Bewegung der Moleküle, bis die Stationärzustandsverteilung erreicht wird.Die umgekehrte TJump und die gleichzeitige Rückdiffusion der fluoreszierenden Moleküle können wie in 1B angegeben beobachtet werden , nachdem der Laser ausgeschaltet ist. Um grundlegende Bindungsparameter, verschiedene Molverhältnisse der Interaktionspartner für den Zugriff werden analysiert und verglichen. Typischerweise werden 16 unterschiedliche Verhältnisse in einem MST Experiment untersucht, wohingegen der optische sichtbaren Molekül konstant gehalten wird und mit zunehmender Menge des unmarkierten Liganden zugeführt wird. Die Wechselwirkung zwischen den beiden Bindungspartner induziert Veränderungen in der Thermophorese, und somit in der normalisierten Fluoreszenz, F Norm, die wie folgt berechnet wird:
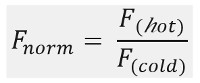
F heiß und F kalt stellen Intensitäten fluoreszenz bei de fi nierten Zeitpunkte der MST Spuren gemittelt. Bindungsaffinitäten (K d oder EC 50 -Werte) können durch curv berechnet werdene Fitting (1C).
Insgesamt ist MST ein leistungsfähiges Werkzeug molekularen Wechselwirkungen jeglicher Art zu studieren. Dieses Manuskript bietet ein Protokoll das herausfordernde Interaktion zwischen dem kleinen Molekül Adenosintriphosphat (ATP; 0,5 kDa) zu charakterisieren und die 25-nt kurze ssDNA Aptamer DH25.42 (7,9 kDa). Im Laufe des Manuskripts wird die Bindungsstelle des Aptamers an das ATP-Molekül an die Adenin-Gruppe des ATP kartiert unten.
