La interacción entre las moléculas es la base de la naturaleza. Por lo tanto, los científicos en muchos campos de la investigación básica y aplicada tratan de comprender los principios fundamentales de las interacciones moleculares de diferentes tipos. MicroScale termoforesis (MST) permite a los científicos para llevar a cabo la caracterización rápida, precisa y rentable, y con control de calidad de las interacciones moleculares en solución, con una libre elección de tampones. Ya hay más de 1.000 publicaciones utilizando MST, a partir de 2016 solo, que describe diferentes tipos de análisis, incluyendo proyecciones de la biblioteca, validaciones de eventos, ensayos de competición, y los experimentos de unión con múltiples parejas de unión 1-8. En general, MST permite el estudio de los parámetros de unión clásicos, tales como la afinidad de unión (pM a mM), estequiometría, y la termodinámica, de cualquier tipo de interacción molecular. Una gran ventaja de MST es la capacidad para estudiar eventos de unión independientes del tamaño de las parejas de interacción. incluso chalinteracciones lenging entre pequeños aptámeros de ácido nucleico (15-30 nt) y objetivos tales como moléculas pequeñas, fármacos, antibióticos, o metabolitos pueden ser cuantificados.
Las tecnologías actuales del estado de la técnica para caracterizar las interacciones aptámero-objetivo son ya sea en laboratorio intensa y de gran complejidad o no se cuantifica aptámero-molécula pequeña Interacciones de 9,10. Surface Plasmon Resonance (SPR) a base de ensayos colorimétricos 11,12 y enfoques verdaderamente libre de marca, tales como calorimetría de titulación isotérmica (ITC) 13-15, elución isocrática 16, el equilibrio filtración 17,18, el sondeo en línea 19, de gel cambiar ensayos, stopped- flujo espectroscopía de fluorescencia 20,21, anisotropía de fluorescencia (FA) 22,23, de una sola molécula de fluorescencia de imagen 24,25, y Bio-capa de interferometría (BLI) 26 también son bien imprecisa o incompatible con la molécula del aptámero microcítico interacciones. otros principal problemas de estos métodos son de baja sensibilidad, alto consumo de ejemplo, la inmovilización, las limitaciones de transporte de masa en las superficies, y / o restricciones de amortiguamiento. Sólo unas pocas de estas tecnologías proporcionan controles integrados para la agregación y adsorción efectos.
MST representa una poderosa herramienta para los científicos para superar esta limitación para estudiar las interacciones entre los aptámeros y moléculas pequeñas 27-29, así como otros objetivos, tales como proteínas 30-33. La tecnología se basa en el movimiento de moléculas a través de gradientes de temperatura. Este movimiento dirigido, llamado "termoforesis," depende del tamaño, la carga, y la capa de hidratación de la molécula 34,35. La unión de un ligando a la molécula alterará directamente al menos uno de estos parámetros, lo que resulta en una movilidad termoforético cambiado. Ligandos con tamaños pequeños pueden no tener un impacto considerable en términos de cambio de tamaño de la independiente al estado ligado, pero pueden tener dr Amatic efectos sobre la capa de hidratación y / o carga. Los cambios en el movimiento termoforético de moléculas después de la interacción con la pareja de unión permite la cuantificación de los parámetros de unión básicas 2,7,34,36,37.
Como se muestra en la Figura 1A, el dispositivo MST consta de un láser infrarrojo enfocado sobre la muestra dentro de los capilares de vidrio utilizando la misma óptica como para la detección de fluorescencia. El movimiento termoforético de las proteínas a través de la fluorescencia intrínseca de triptófanos 6 o de un 3,8 pareja de interacción marcado con fluorescencia se puede monitorizar mientras el láser se establece un gradiente de temperatura (? T de 2-6 ° C). La diferencia de temperatura resultante en el espacio,? T, conduce al agotamiento o la acumulación de moléculas en el área de temperatura elevada, que se puede cuantificar por el Soret coeficiente (S T):
g "/>
c caliente representa la concentración en la región calentada, y c es la concentración de frío en la región en frío inicial.
Como se muestra en la Figura 1B, un típico experimento MST produce un perfil de movimiento MST (traza el tiempo), que consta de diferentes fases, que pueden separarse por sus respectivas escalas de tiempo. La fluorescencia inicial se mide en los primeros 5 s en ausencia del gradiente de temperatura para definir la fluorescencia a partir preciso y para comprobar si hay photobleaching o photoenhancement. El salto de temperatura (T-Jump) representa la fase en la que los cambios de fluorescencia antes del movimiento termoforético. Esta disminución inicial de la fluorescencia depende de cambios dependientes de calor de fluoróforo rendimiento cuántico. La fase termoforesis sigue, en el que se alcanza la disminución de fluorescencia (o aumenta) debido al movimiento de las moléculas termoforético hasta la distribución en el estado estacionario.El TJump inversa y retrodifusión concomitante de moléculas fluorescentes fl se pueden observar como se indica en la figura 1B después de que el láser está apagado. Con el fin de acceder a los parámetros básicos de unión, diferentes relaciones molares de los compañeros de interacción son analizados y comparados. Típicamente, 16 relaciones diferentes se estudian en un experimento MST, mientras que la molécula visible óptico se mantiene constante y se suministra con una cantidad creciente de ligando no marcado. La interacción entre las dos parejas de unión induce cambios en la termoforesis, y así en la fluorescencia normalizada, F norma, que se calcula de la siguiente manera:
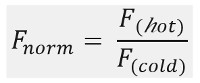
F caliente y fría F representan un promedio de intensidades de fluorescencia en puntos de tiempo de fi nido de las huellas del MST. Las afinidades de unión (Kd o valores de EC50) se pueden calcular por curve apropiado (Figura 1C).
En general, el MST es una poderosa herramienta para estudiar las interacciones moleculares de ningún tipo. Este manuscrito ofrece un protocolo para caracterizar la interacción entre el reto pequeña molécula de trifosfato de adenosina (ATP; 0,5 kDa) y el 25-nt corto ssDNA aptámero DH25.42 (7,9 kDa). En el transcurso del manuscrito, el sitio de unión del aptámero en la molécula de ATP se mapea hasta el grupo adenina del ATP.
