Interaksjon mellom molekylene er grunnlaget for naturen. Derfor forskere i mange felt av grunnleggende og anvendt forskning prøve å forstå de grunnleggende prinsippene i molekylære interaksjoner av ulike slag. Mikro termodiffusjon (MST) gjør det mulig for forskere å utføre raske, presise, kostnadseffektive og kvalitetssikret karakterisering av molekylære interaksjoner i løsning, med et fritt valg av buffere. Det er allerede mer enn 1000 publikasjoner bruke MST, fra 2016 alene, som beskriver ulike typer analyser, inkludert bibliotek screenings, bindende event valideringer, konkurranseanalyser, og eksperimenter med flere bindingspartnere 1-8. Generelt tillater MST studiet av de klassiske bindende parametere, slik som bindingsaffinitet (pM til mM), støkiometri, og termodynamikk, av en hvilken som helst form for molekylær interaksjon. En stor fordel med MST er evnen til å studere bindings hendelser uavhengig av størrelsen på interaksjonspartnere. selv utfordrende interaksjoner mellom små nukleinsyre aptamerer (15-30 nt) og mål som små molekyler, narkotika, antibiotika, eller metabolitter kan kvantifiseres.
Nåværende state-of-the-art teknologi for å karakterisere aptamer-målet interaksjoner er enten lab-intens og svært komplekse eller unnlater å kvantifisere aptamer-lite molekyl interaksjoner 9,10. Surface plasmonresonans (SPR) -baserte analyser 11,12 og virkelig label-free kalori tilnærminger, for eksempel Isoterm Titrering kalorimetri (ITC) 13-15, isokratisk eluering 16, likevekt fi filtrering 17,18, in-line sondering 19, gel- skifte analyser, stopped- fl ow fl uorescence spektroskopi 20,21, fluorescens anisotropi (FA) 22,23, single-molekyl fl uorescence bildebehandling 24,25, og Bio-lag interferometri (BLI) 26 er også enten upresis eller uforenlig med aptamer-lite molekyl interaksjoner. andre Principal utgaver av disse metodene er lav følsomhet, høy prøven forbruk, immobilisering, massetransportbegrensninger på overflater, og / eller buffer restriksjoner. Bare et fåtall av disse teknologiene gir integrerte kontroller for aggregering og adsorpsjon effekter.
MST representerer et kraftig verktøy for forskere å overvinne denne begrensningen for å studere interaksjoner mellom aptamerer og små molekyler 27-29, samt andre mål, for eksempel proteiner 30-33. Teknologien baserer seg på bevegelse av molekyler gjennom temperaturgradienter. Dette rettet bevegelse, kalt "termodiffusjon", avhenger av størrelse, ladning, og hydrering skall av molekylet 34,35. Binding av en ligand til molekylet vil direkte endre i det minste én av disse parametre, noe som resulterer i en endret termoforetiske mobilitet. Ligander med små størrelser kan ikke ha store konsekvenser i forhold til størrelse endring fra ubundet til bundet tilstand, men de kan ha dr amatic effekter på hydrering skall og / eller kostnad. Endringene i den termoforetiske bevegelighet av molekylene etter interaksjon med bindingspartneren muliggjør kvantifisering av grunnleggende bindingsparametere 2,7,34,36,37.
Som vist i figur 1A, MST enheten består av en infrarød laser fokusert på prøven innenfor glasskapillærer ved hjelp av de samme optikk som for fluorescensdeteksjon. Den termo bevegelse av proteiner via den iboende fl uorescence av tryptofaner 6 eller av et fluorescensmerket interaksjon partner 3,8 kan overvåkes mens laseren etablerer en temperaturgradient (aT på 2-6 ° C). Den resulterende temperaturforskjell på plass, AT, fører til uttømming, eller oppsamling av molekyler i området av forhøyet temperatur, som kan kvantifiseres ved Soret COEF fi sient (S T):
g "/>
c varm utgjør den konsentrasjon i det oppvarmede område, og c kulde er konsentrasjonen i den innledende kald region.
Som vist i figur 1B, en typisk MST eksperimentresultatene i en MST bevegelse profil (tid kurve), som består av forskjellige faser, som kan separeres ved deres respektive tidsrammer. Den innledende fluorescensen måles i de første 5 s i fravær av temperaturgradienten for å definere den nøyaktige utgangs fluorescensen og for å se etter fotobleking eller photoenhancement. Temperatur Jump (T-Jump) representerer den fasen hvor fluorescens endringene før termo bevegelse. Denne første nedgang i fluorescens er avhengig av varmeavhengige endringer av fl uorophore kvanteutbytte. Den termodiffusjon fase følger, hvor fluorescens-lavere (eller høyere) på grunn av den termoforetiske bevegelighet av molekylene inntil steady-state fordeling er oppnådd.Det motsatte TJump og samtidig tilbakediffusjon av fl uorescent molekyler kan observeres som vist i figur 1B etter at laseren er skrudd av. For å få tilgang til basisbindingsparametere, blir forskjellige molare forhold av interaksjonspartnere analysert og sammenlignet. Vanligvis er 16 forskjellige forhold studert i en MST eksperiment, mens den optiske synlig molekylet holdes konstant og er utstyrt med en økende mengde av umerket ligand. Samspillet mellom de to bindingspartnere induserer endringer i termodiffusjon, og således i den normaliserte fl uorescence, F norm, som beregnes som følger:
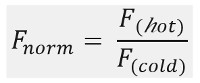
F varm og F kald representerer gjennomsnitt fl uorescence intensiteter på de fi nert tidspunkter av MST spor. Bindingsaffiniteter (K d eller EC 50 verdier) kan beregnes ved curve montering (figur 1C).
Samlet er MST et kraftig verktøy for å studere molekylære interaksjoner av noe slag. Dette manuskriptet har en protokoll for å karakterisere utfordrende samspill mellom den lille molekylet adenosintrifosfat (ATP, 0,5 kDa) og 25-nt kort ssDNA aptamer DH25.42 (7,9 kDa). I løpet av manuskriptet er bindingssetet for aptamer på den ATP-molekylet tilordnet ned til adenin gruppen av ATP.
