The Middle East Asia Minor 1 (MEAM1) species of the B. tabaci complex was taken as an example here for description. Cotton for rearing whiteflies and several developmental stages of whiteflies are shown in Figure 1 including a cotton plant, adult whitefly and the 1st, 2nd and 4th instar nymphs of whitefly (the 3rd instar nymph looks similarly as the 4th instar nymph). It was obvious that the 4th instar nymph is larger than the 1st and 2nd instar nymphs (Figure 1). FISH analysis of Portiera and Hamiltonella within MEAM1 is shown in Figure 2. These two endosymbionts are confined to the bacteriocytes of the whitefly, and overlapping of the two bacteria is also observed. Figure 3 shows transmission electron microscopy images of the Portiera endosymbiont of the whitefly, which indicates that Portiera may lose the its cell wall.
For sequencing, two libraries were built, with insert sizes of 200 bp and 2,000 bp, respectively. After sequencing, the two libraries generated 1,626 Mb and 1,302 Mb of raw data, respectively. After cleaning up the contamination data for adaptors and duplications, 1,484 Mb and 1,219 Mb of clean data were acquired. The assembly successfully resulted in a complete genome sequence for the obligate symbiont, Portiera. In addition, a draft genome of Hamiltonella was also obtained. The assembly based on both 200 bp and 2,000 bp libraries resulted in 138 contigs. According to paired-end relationships, the 138 contigs were further assembled into 89 scaffolds (Table 2).

Figure 1: Overview of the Plant-insect System Utilized in this Paper. (A) Whiteflies are reared on cotton plants (Gossypium hirsutum cv. Zhe-Mian 1793). (B) View of an adult whitefly. (C) Several developmental stages of instar nymph in the whitefly life cycle. Red arrow refers to the egg of whitefly; Black arrow refers to the 1st instar nymph of whitefly; White arrow refers to the 2nd nymph of whitefly; Blue arrow refers to the 4th instar nymph of whitefly. Please click here to view a larger version of this figure.

Figure 2: FISH Analysis of Portiera, Hamiltonella in the 4th Instar Nymph of MEAM1. Portiera-specific probe (red) conjugated to Cy3, Hamiltonella-specific probe (green) marked with Cy5 were used. Scale bars = 100 µm. (A) The red hybridization signals represent the Portiera under dark field. (B) The green hybridization signals represent the Hamiltonella under dark field. (C) Portiera and Hamiltonella merged signals under dark field. (D) Portiera and Hamiltonella merged signals under bright field. Please click here to view a larger version of this figure.
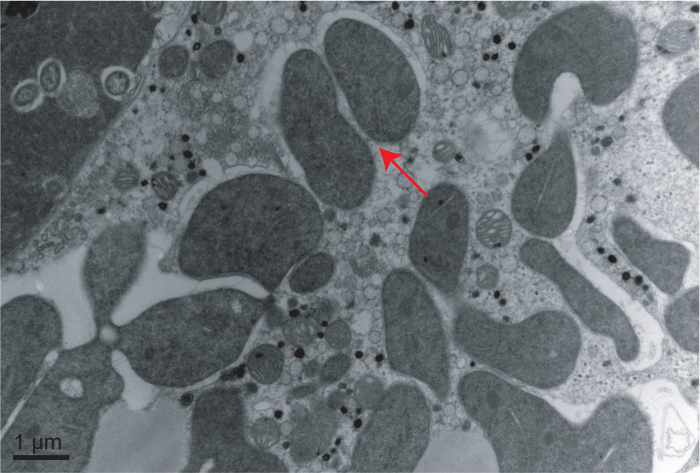
Figure 3: Transmission Electron Microscopy Image of Portiera in Whitefly. Arrow indicates the location of Portiera. Scale bar = 1 µm. Please click here to view a larger version of this figure.
| Target symbiont | Target gene | Primer sequences (5'-3') | PCR procedures | References | ||
| Portiera | 16S rRNA | Por-F: TGCAAGTCGAGCGGCATCAT | 95 °C 1 min, 60 °C 1 min, 72 °C 1 min, 5 cycles; | 27 | ||
| Por-R: AAAGTTCCCGCCTTATGCGT | 95 °C 1 min, 58 °C 1 min, 72 °C 1 min, 30 cycles; | |||||
| 72 °C 20 min | ||||||
| Hamiltonella | 16S rRNA | Ham-F: TGAGTAAAGTCTGGGAATCTGG | 95 °C 1 min, 60 °C 1 min, 72 °C 1 min, 5 cycles; | 27 | ||
| Ham-R: CCCGGGAACGTATTCACCGTAG | 95 °C 1 min, 58 °C 1 min, 72 °C 1 min, 30 cycles; | |||||
| 72 °C 20 min | ||||||
| Rickettsia | 16S rRNA | Ric-F: GCTCAGAACGAACGCTATC | 95 °C 2 min; | 24 | ||
| Ric-R: GAAGGAAAGCATCTCTGC | 92 °C 1 min, 58 °C 1 min, 72 °C 90 s, 30 cycles; | |||||
| 72 °C 5 min | ||||||
| Arsenophonus | 23S rRNA | Ars-F: CGTTTGATGAATTCATAGTCAAA | 95 °C 5 min; | 28 | ||
| Ars-R: GGTCCTCCAGTTAGTGTTACCCAAC | 95 °C 30 s, 60.5 °C 30 s, 72 °C 45 s, 30 cycles; | |||||
| 72 °C 10 min | ||||||
| Cardinium | 16S rRNA | Car-F: TACTGTAAGAATAAGCACCGGC | 95 °C 2 min | 29 | ||
| Car-R: GTGGATCACTTAACGCTTTCG | 92 °C 1 min, 57 °C 1 min, 72 °C 90 s, 30 cycles; | |||||
| 72 °C 5 min | ||||||
| Wolbachia | 16S rRNA | Wol-F: TTGTAGCCTGCTATGGTATAACT | 94 °C 5 min; | 30, 31 | ||
| Wol-R: GAATAGGTATGATTTTCATGT | 94 °C 1 min, 55 °C 1 min, 72 °C 1 min, 35 cycles | |||||
| Fritschea | 23S rRNA | Fri-F: GATGCCTTGGCATTGATAGGCGATGAAGGA | 95 °C 5 min; | 32 | ||
| Fri-R: TGGCTCATCATGCAAAAGGCA | 94 °C 1 min, 64 °C 1 min, 72 °C 90 s, 35 cycles; | |||||
| 72 °C 5 min | ||||||
| Hemipteriphilus | groEL | OR-groEL-F: CACCWAAAATTACTAAAGATGG | 94 °C 3 min; | 18 | ||
| OR-groEL-R: TAGAARTCCATWCCKCCCATWC | 94 °C 30 s, 52 °C 30 s, 72 °C 2 min, 34 cycles; | |||||
| 72 °C 10 min | ||||||
Table 1: PCR Primers and Procedures of Endosymbionts in Whitefly.
| Portiera | Hamiltonella | |
| Contig Number | 1 | 138 |
| Scaffold numbera | 1 | 89 |
| Total length, bp | 358,232 | 1,711,449 |
| N50, bp | / | 118,802 |
| N90, bp | / | 16,753 |
| Max length, bp | / | 390,511 |
| Min length, bp | / | 503 |
| GC content, % | 26.18 | 39.89 |
| a Information presented below are all based on scaffolds. | ||
Table 2: General Features of Portiera and Hamiltonella Draft Genome.
