A schematic of the protocol is provided in Figure 1. The lipoprotein-enriched fraction extracted from Enterococcus faecalis ATCC 19433 by TX-114 is shown in Figure 2. For comparison, the banding pattern of the precipitated protein fraction is also shown. Proteins from this fraction were confirmed by MALDI-MS to be highly abundant contaminating proteins other than lipoproteins (Table 1). The mass spectra in Figure 3 demonstrate the tryptic peptide ion profile of the E. faecalis lipoprotein PnrA that occurs with subsequent washes of nitrocellulose-bound PnrA with solvents of increasing polarity (peak assignments listed in Table 2). Figure 4 shows the N-terminal structural characterization of PnrA as determined by MALDI-TOF MS/MS, revealing diagnostic N-acylated dehydroalanyl peaks consistent with the lyso-lipoprotein form. Figure 5 illustrates the effect of sodium adduct formation on fragmentation, with the sodiated parent ion preferentially fragmenting in favor of the N-acylated dehydroalanyl ion.
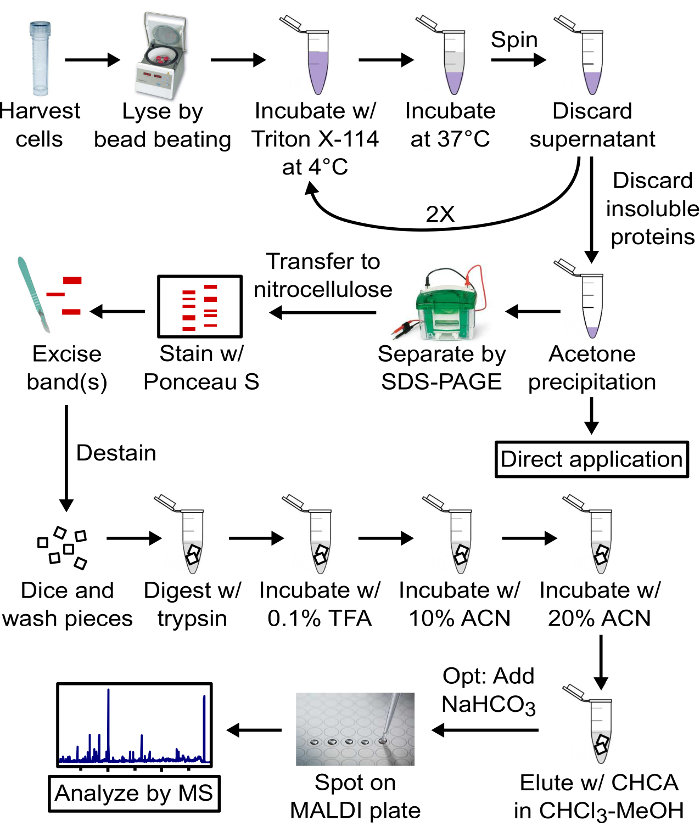
Figure 1: Schematic of the protocol. Lipoproteins are enriched by TX-114 phase partitioning and can be used directly, most commonly in TLR assays, or further purified by SDS-PAGE for structural determination. Lipoproteins are transferred to nitrocellulose, digested with trypsin, washed stepwise, and the resulting lipopeptides eluted with chloroform-methanol for structural analysis by MALDI-MS. The trypsin and nitrocellulose wash solutions may be saved for protein identification and MS analysis. w/: with; TFA: trifluoroacetic acid; ACN: acetonitrile; CHCA: α-cyano-4-hydroxycinnamic acid; opt: optional; MALDI: matrix-assisted laser desorption ionization; MS: mass spectrometry. Please click here to view a larger version of this figure.
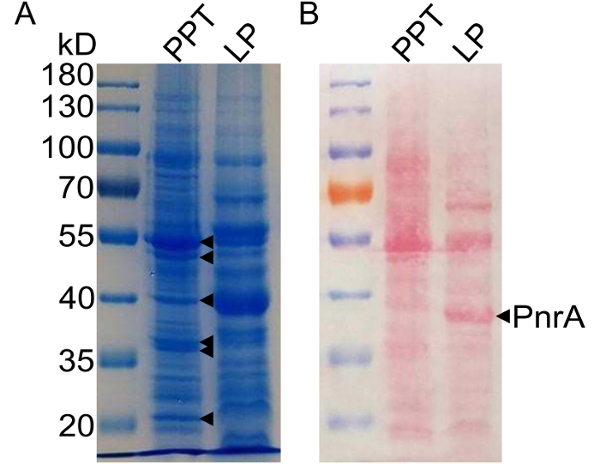
Figure 2: Profile of TX-114 enriched proteins from E. faecalis. A 10% Tris-glycine SDS-PAGE gel stained with Coomassie blue (A) and the corresponding nitrocellulose membrane stained with Ponceau S (B) reveal a different banding pattern between the precipitated protein fraction ("PPT") and the purified lipoproteins ("LP"). The indicated Coomassie-stained bands were excised and identified as non-lipoproteins by MALDI-MS of tryptic peptides (see Table 1). PnrA was identified and analyzed by the protocol described herein. Please click here to view a larger version of this figure.
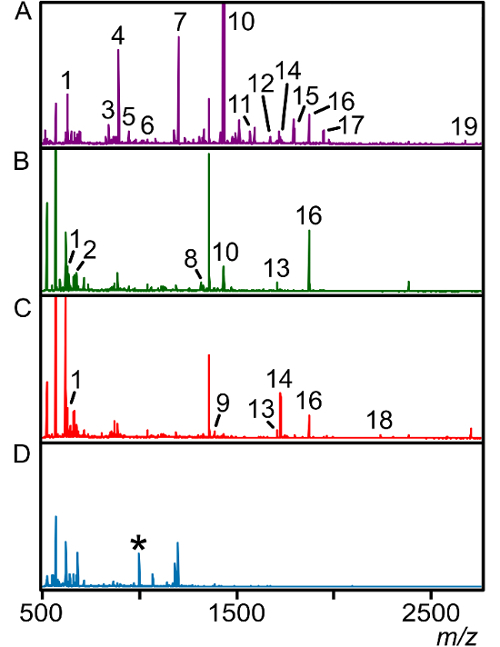
Figure 3: Ion profile and abundance changes with polarity of nitrocellulose wash solutions. (A) The trypsin fraction shows several internal peptide fragments corresponding to the E. faecalis lipoprotein PnrA indicated in Figure 2. The 10% (B) and 20% (C) acetonitrile wash fractions show changes in signal intensity and a decrease in the overall number of individual peaks. (D) The final elution fraction is highly enriched with the N-terminal lipopeptide at m/z 997, indicated by an asterisk (*). Intensity of each spectra is normalized to (A) for comparison of signal intensity. Peaks masses and assigned sequences are listed in Table 2. Please click here to view a larger version of this figure.
| Band | Protein ID | Accession No. | Est. Molecular Weight | Peptide Count | |
| 1 | elongation factor Tu | gb|EOL37301.1| | 43,387 | 15 | |
| 2 | NADH peroxidase | gb|EOL34572.1| | 49,520 | 9 | |
| 3 | pyruvate dehydrogenase E1component, alpha subunit | gb|EOL34709.1| | 41,358 | 12 | |
| 4 | pyruvate dehydrogenase E1 component, subunit beta | gb|EOL34710.1| | 35,373 | 19 | |
| 5 | 30S ribosomal protein S2 | gb|EOL33066.1| | 29,444 | 16 | |
| 6 | 30S ribosomal protein S3 | gb|EOL37312.1| | 24,355 | 14 | |
Table 1: Precipitated proteins are non-lipoprotein contaminants. Proteins were identified by tryptic digest and MALDI-MS using standard in gel digestion protocols19 and are listed from top to bottom in the same order they appear on the Coomassie gel in Figure 2. Each protein was identified with a Confidence Interval (C.I.) greater than 95%.
| Peak Number | Theoretical Mass | Protein Position | No. of Missed Cut Sites | Peptide Sequence | |
| 1 | 631.3409 | 170-176 | 0 | GVADAAK | |
| 2 | 675.4035 | 316-321 | 1 | TKEAVK | |
| 3 | 826.4093 | 163-169 | 0 | FQAGFEK | |
| 4 | 893.4839 | 121-128 | 0 | NVVSATFR | |
| 5 | 943.5359 | 240-247 | 0 | VWVIGVDR | |
| 6 | 1040.5047 | 188-197 | 0 | YAASFADPAK | |
| 7 | 1200.6331 | 273-284 | 0 | GVGTAVQDIANR | |
| 8 | 1316.6997 | 290-301 | 0 | FPGGEHLVYGLK | |
| 9 | 1385.7827 | 83-94 | 0 | FNTIFGIGYLLK | |
| 10 | 1432.743 | 149-162 | 0 | VGFVGGEEGVVIDR | |
| 11 | 1568.7802 | 259-272 | 1 | DGKEDNFTLTSTLK | |
| 12 | 1672.8289 | 240-254 | 1 | VWVIGVDRDQDADGK | |
| 13 | 1707.8184 | 129-145 | 0 | DNEAAYLAGVAAANETK | |
| 14 | 1724.8027 | 35-49 | 0 | SFNQSSWEGLQAWGK | |
| 15 | 1797.9228 | 257-272 | 2 | TKDGKEDNFTLTSTLK | |
| 16 | 1872.9854 | 285-301 | 1 | ALEDKFPGGEHLVYGLK | |
| 17 | 1945.9613 | 230-247 | 1 | DLNESGSGDKVWVIGVDR | |
| 18 | 2240.1345 | 149-169 | 1 | VGFVGGEEGVVIDRFQAGFEK | |
| 19 | 2675.2543 | 230-254 | 2 | DLNESGSGDKVWVIGVDRDQDADGK | |
Table 2: Observed masses and the corresponding tryptic peptides. In silico trypsin digestion of E. faecalis PnrA (GenBank: EOL37280.1) was performed using PeptideMass20,21, allowing up to two missed cut sites. The theoretical masses of the peaks observed on the spectra in Figure 3 are listed, along with the corresponding peptide sequences.
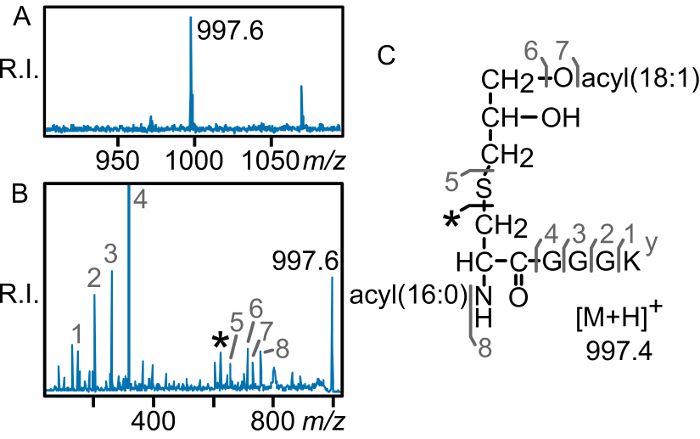
Figure 4: MALDI-TOF MS of the E. faecalis lipoprotein PnrA. (A) Parent MS spectrum of the m/z 997 region corresponding to the N-terminal lipopeptide of E. faecalis PnrA. (B) MS/MS of the lipopeptide peak reveals that it is the lyso form, with the diagnostic N-acylated dehydroalanyl peptide fragment ion peak indicated by an asterisk (*). The elucidated structure is shown (C). This figure has been modified from a previous publication9. R.I.: relative intensity Please click here to view a larger version of this figure.
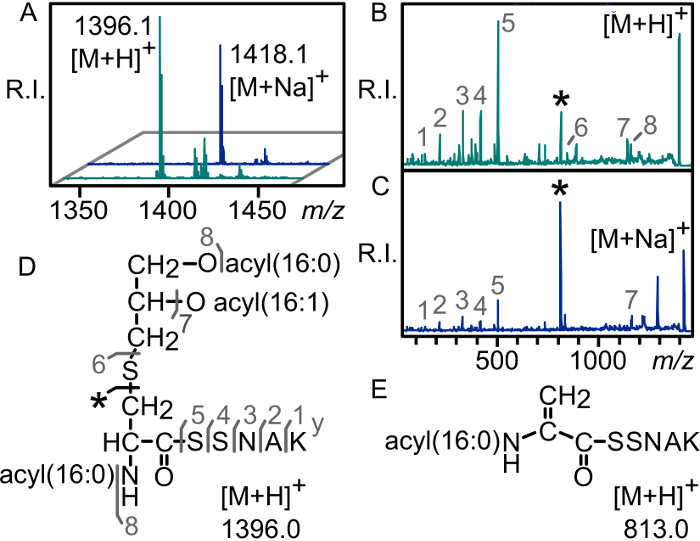
Figure 5: Sodium adduct formation promotes parent ion fragmentation towards dehydroalanyl lipopeptide ions. (A) MALDI-TOF spectra of the E. coli lipoprotein Lpp N-terminal peptide without (turquoise traces) and with (blue traces) the addition of sodium bicarbonate. Addition of sodium bicarbonate to the final eluted fraction results in a 22 Da increase from the calculated mass of the parent lipopeptide. When compared with the MS/MS spectrum of the protonated ion (B), the MS/MS spectrum of the corresponding sodiated ion (C) shows significant preferential fragmentation toward the N-acyl-dehydroalanyl ion through neutral elimination of the diacylthioglyceryl moiety, indicated by an asterisk (*). The structure of the parent triacylated Lpp N-terminal tryptic peptide (D) and the N-acyl-dehydroalanyl peptide fragment ion (E) are depicted. This figure has been modified from a previous publication9. R.I.: relative intensity Please click here to view a larger version of this figure.
