RNA-sequencing analyse van transcriptome lymfklieren biedt de mogelijkheid om het karakteriseren van de immuunrespons van dieren aan een verscheidenheid van pathogenen. Terwijl deze methodologie is gebruikt uitgebreid in muizen, analyses hebben onlangs geweest zich uitbreiden naar grotere zoogdieren1,2. Dieren/grote dierlijke lymfeklieren kan worden gebruikt voor het karakteriseren van host reacties op een infectie, niet alleen voor hun gebruik in het vaccin of genetische studies en voor de identificatie van doelen voor Geneesmiddelenontwikkeling, maar ook als modelsystemen voor menselijke studies op zoönotische ziekten. Bijvoorbeeld in het geval van brucellose (een zoönotische bacteriële ziekte dat effecten een half miljoen mensen over de hele wereld elk jaar), ondanks aanzienlijk hogere kosten, studies in schapen of geiten meer relevant zijn voor de menselijke infectie en menselijk vaccin ontwikkeling dan laboratorium diermodellen. Infectie Muismodellen recapituleren de reticuloendotheliaal systeem infectie maar niet de karakteristieke klinische symptomen3.
In grote dierproeven ten opzichte van dierproeven laboratorium bestaat het proces van weefsel oogsten noodzakelijkerwijs uit een langere vertraging tussen de euthanasie en de weefsel-collectie, die een potentiële uitdaging voor het behoud van kwalitatief hoogwaardige RNA. Intact RNA is essentieel voor de generatie van biologisch relevante transcriptomic gegevens. De generatie van kwalitatief hoogwaardige RNA van weefselmonsters is bijzonder kritisch voor grote dieren pathogen studies uitgevoerd in insluiting faciliteiten. Dergelijke studies zijn inherent moeilijker uit te voeren zoals ze niet alleen goedgekeurde voorzieningen en hoogst opgeleid personeel vereisen, maar ook aanzienlijke financiële kosten, die, afhankelijk van het werk voeren, van tientallen tot honderden duizenden dollars variëren kunnen. Deze soorten studies ook betrekking hebben op een interdisciplinaire samenwerking en multidisciplinaire kennis ten behoeve van hun voltooiing, toe te voegen aan hun complexiteit. Daarom, training op, ontwikkeling van en aanhankelijkheid aan een gestroomlijnd systeem voor de sample collectie en het behoud biedt aanzienlijke voordelen voor downstream moleculaire studies van weefsels van besmette dieren.
De collectie van grotere lymfklieren presenteert extra uitdagingen voor de collectie van weefsel ten opzichte van de vergelijkbare bemonstering van lymfkliertest lymfklieren. De voorbereiding voor de steekproef besnijdenis vereist een basiskennis van de anatomie van de lymfeklier, met inbegrip van de relevante interne structuren. De structuur van een lymfeklier bestaat uit lymfoïde melkklieren omgeven door sinussen gevuld met lymfe. Deze structuren zijn ingesloten in een harde, vezelige capsule. 4 een lymfoïde lobule is de “fundamentele anatomische en functionele eenheid van de lymfeklier” en bestaat uit een diep corticale eenheid, en Wallenberg koorden, follikels en sinussen4 (figuur 1A). B en T-lymfocyten zijn de thuisbasis van de follikels en diep corticale eenheden, respectievelijk. Deze structuren bieden een 3D steiger en faciliteren van de interactie tussen de lymfocyten en antigeen of antigeen presentatie van cellen.
Grove, follikels en diep corticale eenheden kunnen worden geïdentificeerd op het snijvlak geconstateerd als ze een dichtere reticulaire gevlochten bevatten en donkerder dan de sinussen, die bestaan uit een meer delicate reticulaire gevlochten en lichter weergegeven (figuur 1B). Door conventie verwijzen pathologen naar de regio’s van de lymfeknopen zoals de oppervlakkige cortex (follikels), de paracortex (diep corticale eenheden) en de medulla (Wallenberg koorden en sinussen). Een nauwkeurig onderzoek van de drie gewesten werd geacht als beste praktijk routine pathologisch onderzoek richtsnoeren voor lymfklieren5. Merk op dat er een aanzienlijke variatie in de consistentie, grootte en kleur van de lymfeknopen, zelfs binnen een enkel dier. Naarmate de dieren ouder, hun lymfklieren zal meestal kleiner en worden steviger dan die van jongere dieren, meestal als gevolg van een toename van hun bindweefsel en een vermindering van de normale lymfoïde structuur6,7.
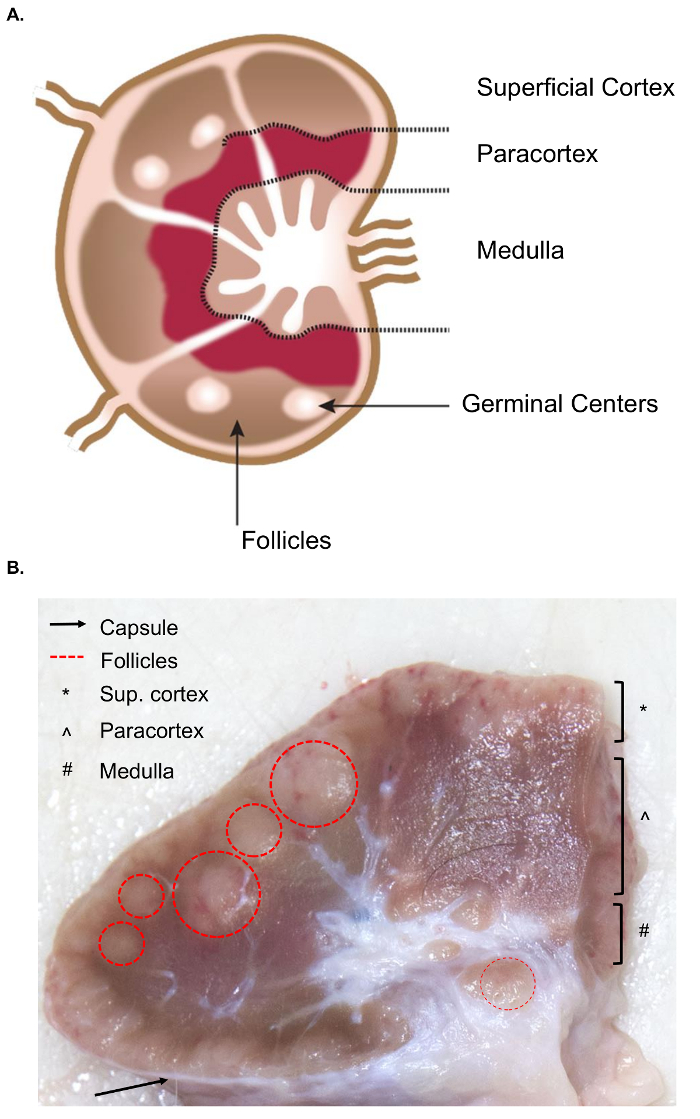
Figuur 1. Anatomie van de lymfeklier. (A) deze cartoon afbeelding toont de anatomie van de lymfeklier, beeltenis van belangrijke structuren. (B) dit nog beeld toont een boviene lymfeklier gesneden in doorsnede. De relevante structuren/lagen die zichtbaar met het blote oog zijn worden gemarkeerd. Klik hier voor een grotere versie van dit cijfer.
Afhankelijk van de experimentele vraag zullen verschillende lymfklieren van belang zijn voor het verzamelen en analyseren. Perifere lymfklieren zijn gelegen diep in het onderhuidse weefsel. Bij runderen, perifere of oppervlakkige lymfklieren vaak gebruikt in de klinische en experimentele praktijk omvatten parotide, submandibulaire retrofaryngeale, prescapulares, prefemoral (precrural) en de oppervlakkige inguïnale (uier worden bij vrouwtjes, scrotum bij mannen) () Figuur 2). In tabel 1, worden de eigenschappen van de belangrijkste oppervlakkige lymfklieren, zoals beschreven in de vee-systeem8, schetste. Hieronder worden enkele potentiële lymfeklier collectie plannen voor bacteriële infectieziekten van runderen gepresenteerd als een uitgangspunt voor het onderzoek.
Brucella abortus/Brucella melitensis: standaard necropsies voor B. abortus-besmet vee en B. melitensis-besmette geiten aan het National dier ziekte Center herstellen uier worden, prescapular en oorspeeksellymfklieren lymfeklier weefsel , zowel voor het slijpen voor de bacteriële opsomming alsmede voor de bereiding van het RNA voor de host RNA expressie profilering. B. abortus kunnen regelmatig worden hersteld in elk van deze lymfeklieren in experimenteel geïnfecteerde runderen9. De aanwezigheid van bacteriën in elk van deze typen lymfeklier kan worden opgespoord in B. melitensis-besmet geiten tot ten minste negen maanden na infectie met behulp van de methoden voor RNA gebaseerde uit onze studies (Boggiatto et al., onuitgegeven). Salmonella sp.: de prescapular, subiliac (prefemorale), en de mesenterische lymfklieren zinvol zijn geweest tijdens de profilering van runderkarkassen voor een Salmonella prevalentie10,11,12 en zou van potentieel belang voor transcriptomic studies. E. coli O157:H7: mesenterische lymfklieren (op de middelste dunne darm en distale dunne darm locaties) kunnen de sites van een occasionele herstel van de bacteriën in geïnfecteerde kalveren (maar niet in geïnfecteerde volwassen runderen)13. Leptospirose (Leptospira sp.): een chronische persistentie van de bacteriën is waargenomen in de lymfeknopen aftappen van de melkklier14. Mycobacterium bovis : Bij runderen, zijn de bacteriën herstelde na experimentele infectie van de lymfeknopen mediastinale en tracheobronchial van kalveren15geweest. Bovendien is lymfeklier RNA te onderzoeken dieren talloze reacties op virussen, zoals de varkens reproductieve en respiratoir syndroom virus2gebruikt. Figuur 2 toont de locatie van een subset van deze grote lymfeklieren in het lichaam van vee.
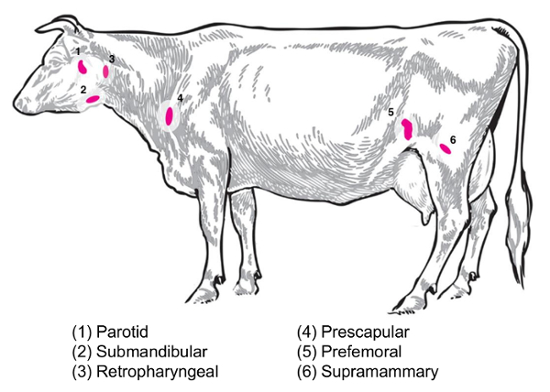
Figuur 2: Cartoon beeltenis van geselecteerde lymfeklier locaties in Bos taurus . De genummerde lymfeklieren zijn geannoteerd. Klik hier voor een grotere versie van dit cijfer.
In dit document en de bijbehorende video presenteren wij een protocol voor de isolatie van grote dieren lymfklieren voor RNA studies, ontworpen als informatief voor moleculaire biologen die betrokken zijn bij transcriptomic studies van grote dierlijke infecties. Eerst, wij bieden een overzicht van de isolatie-procedure voor de lymfeknopen, met behulp van bemonstering van weefsels van runderen en bison als voorbeelden. Gekoppeld aan deze demonstratie, zoals weergegeven in de video, is een werkstroom voor een reproduceerbare weefsel bemonstering voor RNA isolatie. Vervolgens beschrijven we belangrijke overwegingen voor de verwerking van een geïnfecteerde lymfe-knooppunt, met een focus op veiligheid, consistentie en kwaliteit van RNA.
De voorbereiding van RNA van het weefsel met een aangezuurde fenol-guanidine isothiocyanaat reagens is gebaseerd op de originele methode van Chomczynski en Sacchi16,17, met een zuivering over silica gebaseerde spin kolommen in aanwezigheid van chaotropic agenten op basis van het oorspronkelijke werk van Vogelstein en Gillespie18. We onderzoeken ook de mogelijkheden voor het herstel van RNA voor transcriptomics van vee lymfklieren geconserveerd door toepassing van alternatieve methoden. Ten slotte onderzoeken we de invloed van de variabele van de tijd op de kwaliteit van de RNA in grote dierlijke necropsies, met inbegrip van een representatieve experiment beeltenis van het effect van een toename van de tijd tussen de euthanasie en de bemonstering op de herstelde RNA-Profiel van bison en boviene lymfklieren. Dit artikel zal nuttig zijn niet alleen voor moleculaire biologen maar ook aan veterinaire onderzoekers vanaf transcriptomic studies zijn.
