Dissociationskonstant K är en viktig parameter att beskriva en receptor (R) affinitet för dess ligand (L). Baserat på lagen i massa åtgärder, definieras K för equilibriumen, där receptor-ligand komplexa RL dissocierar i receptorn R och liganden L:
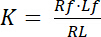 Ekvation 1
Ekvation 1
med index f som visar gratis/unbound delstaten receptor och ligand. Koncentrationen av receptor-ligand komplexet, RL, är identisk med koncentrationen av receptor-bundna ligand Lb. Som den totala koncentrationen av receptorn Rt är summan av den gratis receptorn Rf och ligand-bundna receptorn Rb = Lb, dissociationskonstant kan också skrivas som:
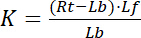 Ekvation 2
Ekvation 2
Därför mättnad avkastning Y, definierad som andelen bundna ligand Lb i förhållandet av den totala koncentrationen av receptorn Rt,
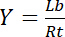 Ekvation 3
Ekvation 3
beror på koncentrationen av fria liganden Lf:
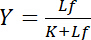 Ekvation 4
Ekvation 4
Denna hyperbolisk beskriver bindande kurvan för en receptor-ligand interaktion och sin tomt visar koncentrationen av bundna ligand Lb som funktion av koncentrationen av fria liganden Lf. Från kurvan är bindande, kan dissociationskonstant K härledas som koncentrationen av fria liganden på halv maximal mättnad yield. Dessutom har olika algoritmer till linjär bindande kurvor fastställts, såsom dubbel-ömsesidiga tomten av Klotz1,2, eller transformationer enligt mikrosatellitsmarkörer eller Hanes (granskas av Bisswanger3). Dock lider alla algoritmer av problem som det högsta värdet av mättnad avkastningen, som att närma sig asymptotiskt vid höga koncentrationer av gratis ligand i bindande kurvan, har uppskattas i en grafisk före utvärdering och därför är fel-benägen.
Dessutom, kräver fastställande av en bindande kurva kvantifiering av fria och bundna ligand under bindande equilibriumen. För detta ändamål har den fria liganden avskiljas från den receptor-bundna liganden och kvantifieras. Därför har den ligand och receptor skiljer sig i deras egenskaper, till exempel en icke-protein ligand i motsats till en protein receptor. Om parterna bindande proteiner, måste de vara distinguishable i deras storlek, avgifter eller andra molekylär funktioner. Kvantifiering av liganden koncentrationer i småskaliga bindande metoder är dock en svår uppgift. Radioaktiv märkning av liganden har ofta varit nödvändig för att upptäcka den låga koncentrationen av bundna ligand, särskilt om stora mängder receptorer inte var tillgängliga eller prisvärda. Dessutom kan den receptor-bundna liganden separera under och efter isolering i en icke försumbar sätt. Komplexa metoder, såsom jämvikt gel filtrering4, kapillärelektrofores5och puls proteolys6, krävs därför att kvantifiera receptor-bundna ligand och skilja det från gratis ligand.
I motsats till dessa bindande analyser kräver titrering experiment inte kvantitativa separation av bundna och fria liganden. I detta syfte titreras en receptor på en konstant koncentration med olika koncentrationer av extra ligand. Genom bindning till receptorn har den bundna liganden biofysiska egenskapen som skiljer den från den fria liganden och är mätbara genom, t.ex., fotometri, fluorometry eller påvisande av antikroppar. Således en signal S, som är proportionell till mättnad avkastning Y och följaktligen också i koncentrationen av receptor-bundna ligand (Lb), identifieras som en funktion av den totala koncentrationen av extra ligand (Lt). Både parametrar, signalen S och den totala koncentrationen av extra ligand är kvantifierade på ett direkt och enklare sätt än koncentrationerna av bundna och fria liganden. Särskilt, tillåtna detektion av receptor-bundna ligand av enzyme-linked immunosorbent assay (ELISA) minskning av provvolymer till under 100 µL samt parallella mätningar av flera ligand koncentrationer i flera mikrotiter plattor. I en titrering ELISA, är en receptor fysiskt adsorberas till en mikrotiterplattan i samma koncentration och titreras med lösliga ligand. Receptorn är orörlig till plastytan i huvudsak genom hydrofob adsorption. Yta koncentrationen av immobiliserade receptor korrelerar med beläggning koncentrationen av receptorn i en ickelinjär relation, sannolikt enligt Langmuir´s adsorption isotherm7. Förutom det totala antalet adsorberade mottagarmolekyler är deras aktivitetsläge en annan viktig parameter för titrering analyser. Endast orörlig receptorer som har behållit ligand bindande aktivitet, är relevanta för titrering analysen och så småningom bidra till den totala koncentrationen av aktiva receptorer Rt titrering analysens, som inte kan fastställas direkt.
Platser på plastytan, som inte omfattas av immobiliserade receptorn är benägna att adsorbera andra proteiner, såsom liganden. Fysiska adsorption av liganden till sådan plast yta webbplatser skulle resultera i en liknande signal som den receptor-bundna liganden, men på ett sätt som ospecifik. Att minska denna ospecifik signal, plast yta platserna av de mikrotiter plattor som inte har varit belagd med protein men blockeras med bovint serumalbumin (BSA). Dock för vissa receptor-ligand titrering analyser, kan ospecifik bakgrund signaler observeras. Då, andra blockerande medel, såsom en lösning på 0,2% gelatin eller 0,04% Tween 20, rekommenderas.
Efter bindning till receptorn avlägsnas den fria liganden av två tvätt steg. Bundna ligand förblir med receptorn, som är orörlig på plast ytan av mikrotiter brunnen, och eventuellt förstärkt av kemiska fixering. För efterföljande kovalent cross-sammanlänkningen av bundna ligand och immobiliserade receptor med glutaraldehyd, buffert ämnet TRIS ersätts för HEPES, utan någon förändring i liganden bindande. HEPES, i motsats till TRIS, inte inaktivera glutaraldehyd. Kovalent cross-länkaget med glutaraldehyd fixar den bundna liganden med dess receptor och förhindrar dess dissociation under efterföljande tvättning och inkubation steg. Således, receptor-ligand interaktionen är kemiskt fryst och garanterar en Titrerkurvan som är opåverkad av efterföljande steg tvätt och inkubation. Glutaraldehyd fixering kan dock kemiskt ändra ligand och receptor på ett sådant sätt att deras interaktion är sänktes eller avskaffades. Ändring av epitoper inom liganden kan dessutom ändra bindande affinitet upptäcka antikroppen, särskilt om en monoklonal antikropp används för att kvantifiera bundna ligand. Även om ingen av dessa negativa effekter av glutaraldehyd fixering sker i denna titrering ELISA, måste känsligheten hos testet mot glutaraldehyd testas för varje receptor-ligand interaktion före titrering experimentet. Efter fixering avlägsnas överflödigt glutaraldehyd i tre tvätt steg med innehållande TRIS buffert. TRIS inactivates återstående aldehyd-grupper, som nonspecifically kan reagera med påvisande av antikroppar i efterföljande steg.
Mängden bundna ligand kvantifieras med enzymkopplad antikroppar, som ger en fotometriska ELISA signal S. Detta är plottade kontra den totala ligand koncentration Lt lagt till varje brunn. Trots dess lättare förvärv är Titrerkurvan inte en hyperbolisk funktion i motsats till bindande kurvan. Dessutom att det har varit oklart hur man beräknar dissociationskonstant K från en Titrerkurvan. Även om algoritmer till linjär spectroscopically förvärvade titrering kurvor har rapporterats självständigt av Stockell8 och Heyn och Weischet9, de föll kort på grund av deras osäkerhet för att beräkna den maximala signalen värde som den mättnad avkastning metoder vid höga koncentrationer av extra ligand.
Här, som en titrering ELISA och en icke-linjär regression algoritm beskrivs att härleda dissociationskonstant K för en receptor ligand interaktion från en Titrerkurvan. Detta protokoll exemplifieras för samspelet mellan kollagen bindande A-domänen för den integrin α2β1 med en snake venom-derived hämmare. Integriner är Celladhesionmolekylar, som medlar ankringen av celler till omgivande extracellulärmatrix eller underliggande basalmembranet10,11. Dessutom förmedla integriner viktiga signaler mellan celler och den extracellular matrisen genom att rekrytera ytterligare signalmolekyler och bildar nya cell organeller, adhesomes, på cell-matrix interaktion12,13, 14. kollagen, liganden av α2β1-integrin, är det mest förekommande proteinet i den mänskliga kroppen och är en avgörande byggnadsställningar komponent av bindväv15. Samspelet mellan α2β1-integrin och kollagen medieras av A-domänen för den integrin α2 subuniten. Integrin α2A-domänen innehåller en tvåvärda cation, som krävs för kollagen bindande och stabiliserar dess struktur. Såväl vildtyp form som mutanter av α2A domänen, till exempel där ytan-utsatt återstoden Y216 hade ersatts för en glycin, kan enkelt recombinantly som produceras i en bakteriell uttryck systemet och isolerade via deras oligo-hans-Taggar med en NiNTA Superflow kolumn med en efterföljande dialys mot TRIS-buffrad koksaltlösning (TBS; 50 mM TRIS/HCl, pH 7,4, 150 mM NaCl) innehållande 2 mM MgCl216. Deras koncentrationer bestämdes med bicinchoninic syra analys (BCA) och deras föroreningar är testade av konventionella SDS-PAGE och fläckade Coomassie-lysande blå R250.
Samspelet mellan α2β1-integrin och kollagen blockeras genom bindning av komponenten venom snake, rhodocetin, från Malayan pit viper (Calloselasma rhodostoma)16,17. Används som en löslig ligand i denna titrering ELISA, rhodocetin renades från rå venom som tidigare beskrivits16. Det löses i HEPES-buffrad koksaltlösning (HBS; 10 mM HEPES/NaOH, pH 7,4, 150 mM NaCl) och förvaras fryst vid-20 ° C. Dess koncentration bestämdes av BCA och dess renhet bevisades av SDS-PAGE. Som en antagonist, rhodocetin inte bara blockerar kollagen bindande till integrin α2β1 A-domän, men också stabiliserar inaktiva konformation av integrin vilket hindrar någon signalering från kollagen i blodkroppar eller blodplättar18. Det är av stor biomedicinsk betydelse Bestäm konstanten dissociation av rhodocetin med dess receptor mål och därmed avslöja dess molekylära mekanism och farmaceutiska potentiella t.ex., som ett antitrombosmedel. För detta ändamål, en titrering ELISA beskrivs inklusive dess utvärdering, som är tillämpligt till nästan alla receptor-ligand interaktion med en 1:1 interaktion stökiometri.
