نقص، أي حل تركيزات الأكسجين [] بما فيه الكفاية منخفضة تؤثر تأثيراً سلبيا على البيولوجية والعمليات الإيكولوجية ولكن في كثير من الأحيان وظيفيا ويعرف [القيام] < 2 مغ/لتر1، واكسجين (معرف وظيفيا ك [دو] من 0.0 0.2 مغ/L) تحدث أكثر تواترا وشدة في المياه الساحلية في العالم، ومصبات الأنهار و أعماق المحيطات2،3 ، وكثيراً ما تتفاقم بزيادة المغذيات4،5. مع حد مساحية متزايد من نقص واكسجين، تتأثر سلبا على الفونة العيانية وفقدان الموئل مدى ونوعية الموئل. ومن المتوقع تغير المناخ إلى تفاقم نقص واكسجين6.
في مصبات الأنهار الطبقية، وإثراء المغذيات كثيرة مثل خليج تشيسابيك، الولايات المتحدة الأمريكية، يمكن أن يسود نقص موسميا المستمرة ويمكن أن يحدث سنة بعد2. وباﻹضافة إلى ذلك، ركوب الدراجات البحرية من نقص تواترا في مصبات الأنهار مثل خليج تشيسابيك ومواقع أخرى ويحدث متأخراً أثناء الليل أو في ساعات الصباح الباكر في الصيف7،8.
وركزت معظم الدراسات على أثر التعرض المستمر للكائنات الحية إلى منخفضة [دو] وتسامحهم لنقص واكسجين9،،من1011،،من1213،14 . وعلاوة على ذلك، بحثت دراسات في التحول على نطاق واسع في تكوين الأنواع التوزيعات ووفرة الأنواع استجابة لتمديد منخفضة [دو]4،15. وكثيراً ما الأنواع التي حساسة جداً منخفضة [،] يموت في الجماهير،16 تحويل الأنواع المتبقية حيوانات الأصغر، أصغر حجماً، ولم تدم طويلاً كما، على سبيل المثال، وجدت في النظام الإيكولوجي “الجرف” تكساس لويزيانا4.
عادة ما تسبق التغيرات السلوكية انهيار المجتمع17 وأفادت الدراسات على الاستجابات السلوكية للكائنات الحية الموسعة منخفضة [دو]4،16،،من1718،19 ،،من2021،،من2223،24،25. ومع ذلك، هذه الدراسات، ولا تركز على الردود الواردة من الكائنات الحية لركوب الدراجات البحرية التعرض لنقص والطبيعة المتقلبة من توافر [] في مصبات الأنهار.
نقص ركوب الدراجات البحرية في مصبات الأنهار الضحلة تلقت الوعي المتزايد كما رصد الدراسات [] أكثر غالباً على مدى الأيام مع بارامترات في مصبات الأنهار16،26. يمكن أن تبقى المياه التاكسج لساعات في نهاية الليل أو ساعات الصباح الأولى في فصل الصيف عندما يكون هناك لا عملية التمثيل الضوئي توليد الأكسجين خلال الليل لكن تستهلك الأوكسجين عالية التنفس الهوائي7،16. ووجد أيضا أن أثر المد والجزر البحرية ركوب الدراجات منخفضة الظروف تفعل مع معظم المدقع الدنيا لاحظ عند المد والجزر المنخفضة وتزامنت مع نهاية ليلة27. إلا بعد عدة ساعات من نقص [القيام] يعود إلى نورموكسيا7،،من1628 في دورة يومية.
لتحديد استجابة فيرجينيكا جيم- السلوكية لنقص ركوب الدراجات البحرية والأس الهيدروجيني ونحن رصد فتح وإغلاق الصمامات المحار يتعرضون للمختبرات التي يسببها البحرية ركوب [دو] والدورية الأس الهيدروجيني. وقد استخدمت تثاءب ردود ذات الصدفتين للكشف عن الظروف البيئية المعاكسة. صمام إغلاق ذات الصدفتين في استجابة للملوثات29،،من3031، الطحالب السامة32،،من3334، والتلوث الحراري35،36 , 37، تقليل كمية الغذاء38،،من3940، تغذية نظام39،41، وانبثاق37،42، كبيرة43 , تم قياس 44والأس الهيدروجيني45،46، والجمع بين الرقم الهيدروجيني والأوكسجين الذائب47 . تقنيات تثاءب، على سبيل المثال، شملت الملاحظات المباشرة48،49،13، قياسات مستمر باستخدام مفاتيح ريد والمغناطيس (مراقبة بلح)50، أو مجسات الألياف الضوئية 51 التي تتطلب المياه واضحة. وبالإضافة إلى ذلك، استخدمت أجهزة استشعار قاعة قوة المغناطيس والحقل المغناطيسي لدراسة بلح البحر تثاءب زاوية52،53،،من5455، ونظام الحث كهرومغناطيسي عالية التردد ويمكن قياس المسافة متفاوتة بين اثنين من لفائف الكهربائية التي يتم لصقها في الصمامات استخدام56،57،،من5859. مصدر جهد العالي مطلوب للنظام بالحث الكهرومغناطيسي والسلطة يجب أن يتم تسليمها إلى كلا الجانبين من شل52. هذا النظام أيضا متوفرة تجارياً ك “موسيلمونيتور” (http://mosselmonitor.nl/).
على ميزانية ضيقة لبحث، ونحن شيدت جهاز قياس الضغط غير مكلفة (SGM) استمرار لقياس تثاءب المحار عبر البحرية الناجمة عن معمل الدراجات [دو] ودرجة الحموضة، تحت ظروف الرؤية المنخفضة. نظامنا أيضا أبسط بكثير من نظم المتنافسة، مما يسمح ليتم تجهيزها من خلال تجربة العديد من الحيوانات. أردنا تحديد استجابات فيرجينيكا جيم- السلوكية للبحرية ركوب الدراجات الهوائية الشديدة ([] = 0.6 مغ/لتر) نقص مع مراقبة درجة الحموضة (الأس الهيدروجيني = 7.8) وركوب الأس الهيدروجيني (pH = 7.8-7.0)، على التوالي، وتثاءب الردود إلى معتدل ([] = 1.7 مغ/لتر) نقص. وعلاوة على ذلك، كنا نرغب في تحديد ما إذا كانت قادرة على الاستجابة بسرعة للتغيرات في [] على البحرية دورة وكيفية الاستجابة عندما يعود نورموكسيا بعد حدث التاكسج المحار. ربما المحار يتم تكييفها على النحو الأمثل لسرعة تقلب البيئة التي وجدت في العديد من مصبات الأنهار16،27 التي يعيشون فيها. بينما تتوفر شاشات تثاءب صمام أكثر تعقيداً، يوفر SGM أسلوب غير مكلفة تتيح قياسات مستمرة لصمام تثاءب في المياه حتى في ظروف الرؤية المنخفضة.
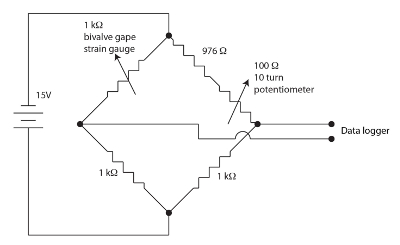
الشكل 1. جسر ويتستون لجهاز صمام تثاءب. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
أجهزة الاستشعار سلالة الاستشعار المستخدمة لرصد تثاءب ذات الصدفتين هي الأفلام مقاوم في نمط تسكع على بوليميد النسخ. كميات صغيرة من سلالة تعدل المقاومة من أجهزة الاستشعار. الصدفتين تنثني سبر سلالة عندما جيبس أنها تسبب تغييرا في مقاومة أجهزة الاستشعار. استخدمنا جسر ويتستون نولينج، ومتوازنة، لكل قناة ذات الصدفتين كما هو مبين في الشكل 1 لقياس التغير في مقاومة أجهزة الاستشعار. جسر ويتستون هو بلا قيمة بمقياس الجهد الانزلاقي السماح مكسب عالية إلى حد ما أن تستخدمها في datalogger. جسر ويتستون أسلوب قياسي لقياس دقة بمقاومة مجهولة باستخدام نسبة إلى مقاومة معروفة قياسية والفولتميتر. وتناقش تاريخ هذه التقنية قديمة جداً في اكيلوف (2001)60. نحن دمج القنوات 12، مع كل جسر ويتستون ومقاومة متغيرة نولينج، وحدة مراقبة قياس الضغط (SGM).
