Hypoxie, dat wil zeggen, ontbonden zuurstofconcentraties [doen] voldoende laag om het negatief beïnvloeden van biologische en ecologische processen zijn, maar vaak functioneel gedefinieerd als [doen] < 2 mg / L1en Anoxie (functioneel omschreven als [] van 0.0-0,2 mg/L) zich voordoen vaker en ernstig in de kustwateren van de wereld, estuaria en de diepe oceaan2,3 en worden vaak verergerd doordat eutrofiëring4,5. Met een toenemende areal omvang van hypoxie en Anoxie, macrofauna ondervinden en verliezen habitat omvang en kwaliteit van habitat. Klimaatverandering wordt voorspeld te verergeren van hypoxie en Anoxie6.
In vele gestratificeerde, nutriënt-verrijkt estuaria zoals Chesapeake Bay, USA, seizoensinvloeden persistente Hypoxie kan zegevieren en jaar na jaar2kan optreden. Bovendien, diel-fietsen van hypoxie is frequent in estuaria zoals de Chesapeake Bay en andere locaties en treedt op tijdens de nacht of de vroege ochtenduren in de zomer7,8laat.
De meeste studies zijn gericht op het effect van continue blootstelling van de organismen tot lage [] en op hun tolerantie voor hypoxie en Anoxie9,10,11,12,13,14 . Bovendien, studies hebben gekeken naar de grootschalige verschuiving in de samenstelling naar soort distributies, abundanties, en soorten in reactie op uitgebreide lage []4,15. Vaak soorten die zeer gevoelig zijn voor laag [doen], sterven in massa,16 verschuiven van de overige soorten om een jonger, kleiner, kortstondige fauna als, bijvoorbeeld, gevonden op de plank van de Louisiana-Texas ecosysteem4.
Gedragsveranderingen meestal ervóór Gemeenschap ineenstorting17 en studies hebben gemeld op gedrags reacties van organismen tot uitgebreide lage []4,16,17,18,19 ,20,21,22,23,24,25. Deze studies, echter, richt u niet op de reacties van organismen diel-Fiet & Mountainbike blootstelling van hypoxie en de wisselende aard van [] beschikbaarheid in estuaria.
Diel-Fiet & Mountainbike hypoxie in ondiepe estuaria heeft ontvangen bewustmaking als studies volgen meer [doen] vaak in de loop van dagen met sondes in estuaria16,26. Water kan blijven hypoxische uren aan het eind van de avond of vroeg in de ochtenduren in de zomer wanneer er geen fotosynthese zuurstof genererende tijdens de nacht, maar hoge zuurstof verbruikt aërobe ademhaling7,16. Ook bleek dat de getijden beïnvloed de diel fietsen voor omstandigheden met weinig doen met de meest extreme minima waargenomen als lage waterstanden viel samen met het einde van de nacht-27. Pas na enkele uren van hypoxie [doet] terug naar normoxia7,16,28 komen in de dagelijkse cyclus.
Om te bepalen van de gedragsmatige reactie van C. virginica diel-Fiet & Mountainbike hypoxie en pH we gecontroleerd het openen en sluiten van de kleppen van oesters blootgesteld aan laboratorium geïnduceerde diel fietsen [do] en cyclische pH. Gape reacties van tweekleppigen zijn gebruikt om het detecteren van negatieve milieu-omstandigheden. Ventiel sluitingen van tweekleppigen in reactie op verontreinigingen29,30,31, giftige algen32,33,34, thermische vervuiling35,36 , 37, voedsel hoeveelheid afnemen38,39,40, voederen regeling39,41, emersion37,42, fotoperiode43 , 44, de pH45,46, en de gecombineerde pH en de opgeloste zuurstof47 zijn gemeten. Gape technieken hebt, bijvoorbeeld directe waarnemingen48,49,13, opgenomen continue metingen met behulp van riet schakelaars en magneten (Dreissena monitor)50, of glasvezel sensoren 51 waarvoor helder water. Daarnaast zijn magneet en magnetische velden veldsterkte Hall sensoren gebruikt om de studie van Mossel gape hoek52,53,,54,55, en een hoge frequentie elektromagnetische inductie systeem dat kan maatregel de variërende afstand tussen twee elektrische spoelen die zijn gelijmd op de kleppen is56,57,58,59gebruikt. Een hoge spanningsbron nodig is voor de elektromagnetische inductie systeem en macht moet worden geleverd aan beide zijden van de shell-52. Dit systeem is ook commercieel beschikbaar als de “MOSSELMONITOR” (http://mosselmonitor.nl/).
Op een strakke onderzoeksbudget, we gebouwd een goedkope spanningsmeter monitor (SGM) voor het continu meten oester gape over laboratorium-geïnduceerde diel fietsen [do] en pH, onder lage zicht. Ons systeem is ook veel eenvoudiger dan concurrerende systemen, waardoor veel dieren worden geïnstrumenteerd tijdens een experiment. We wilden de gedrags reacties van C. virginica op diel fietsen ernstige bepalen ([doen] = 0,6 mg/L) hypoxie met controle pH (pH = 7.8) en fietsen pH (pH = 7.8-7.0), respectievelijk, en gape reacties op milde ([doen] = 1,7 mg/L) hypoxie. Bovendien, we wilden om te bepalen als oesters zijn in staat om snel te reageren op veranderingen in [doen] over de diel cyclus en hoe ze reageren als normoxia na een hypoxische evenement terugkeert. Misschien zijn de oesters optimaal aangepast aan de snel wisselende omgeving die is te vinden in vele estuaria16,27 waar ze wonen. Terwijl de meer complexe ventiel gape monitoren beschikbaar zijn, biedt de SGM een goedkope techniek waarmee continumetingen van ventiel gape in wateren zelfs in lage zicht.
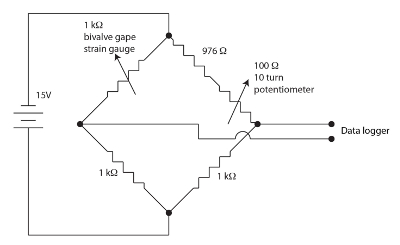
Figuur 1. De brug van Wheatstone voor het ventiel gape apparaat. Klik hier voor een grotere versie van dit cijfer.
De spanningsmeter sensoren gebruikt voor de controle van tweekleppige gape zijn resistieve films in een meander patroon op een polyimide back-ups maken. Kleine hoeveelheden van stam moduleren de weerstand van de sensor. De tweekleppige buigt de strain gage wanneer het gaapt veroorzaken een verandering in de weerstand van de sensor. We gebruikt een nul, evenwichtige, brug van Wheatstone voor elk tweekleppige kanaal zoals afgebeeld in Figuur 1 voor het meten van de verandering in de weerstand van de sensor. De brug van Wheatstone is nul door de potmeter toe te staan van een vrij hoge winst kunnen worden ingezet door de datalogger. Een brug van Wheatstone is een standaardmethode voor het nauwkeurig meten van een onbekende weerstand met behulp van een verhouding tot een bekende weerstand standaard en een voltmeter. De geschiedenis van deze zeer oude techniek wordt besproken in Ekelof (2001)60. We 12 kanalen, elk met een eigen brug van Wheatstone en nul potentiometer, geïntegreerd in de eenheid spanningsmeter Monitor (SGM).
