العظام نوع من النسيج الضام، ويتألف من جزأين: العضوية (خلايا وألياف الكولاجين) والمعدنية (مركبات الكالسيوم والفوسفات). العناصر المعدنية الرئيسية في العظام، أباتيتيس1. أنواع مختلفة من الخلايا المختصة تمعدن في العظام (خلايا الاوستيوبلاستس) والأسنان (أودونتوبلاستس) في الغضروف (تشوندروسيتيس) تنظيم الخطوات الأولية التمعدن بإنتاج بروتينات المصفوفة خارج الخلية (ECM) والإفراج عن مصفوفة حويصلات (MVs) (الشكل 1). MVs هي 100-300 نانومتر القطر الحويصلات التي تتراكم الكالسيوم والفوسفات تيسير التنو الاباتيت وربط الكولاجين2،3في وقت لاحق. ثم، تتفكك MVs للإفراج عن أباتيتيس إلى الوسط خارج الخلية. لا تزال أباتيتيس تنمو على اتصال بألياف الكولاجين وتشكل مصفوفة العظام. التمعدن تدعمه إمدادات ثابتة من فأنا و Ca2 + في الوسط خارج الخلية. بعض البيانات المنشورة مؤخرا الدعم لدينا نموذج4،5. لا طاغية الأنسجة الرخوة تحت الظروف الفسيولوجية. ومع ذلك، قد يحدث تكلس حمل خارج الرحم تحت الظروف المرضية مثل تكلس الأوعية الدموية3. يمكن أن تنتج خلايا الأوعية الدموية التي تحصل على النمط الظاهري osteoblast MVs حمل نويات أباتيتيس والشروع في تمعدن في طبقات الجدار الآنسي وإينتيمال من الأوعية الدموية. منذ تكلس حمل خارج الرحم تشبه endochondral العادي التمعدن3، فهم الآليات الجزيئية التمعدن العظمى الخلايا وتشوندروسيتيس ينبغي أن توفر بعض القرائن على حمل خارج الرحم تكلس الأنسجة الرخوة التي شكلت.
وينظم وضع أنسجة الهيكل العظمى الإنزيمات المختلفة، عوامل النمو، والمروجين أو مثبطات التمعدن. إجراءات معادية للأنسجة-غير محددة الفوسفاتيز القلوية (طنب) (الشكل 1) واكتونوكليوتيدي بيروفوسفاتاسي/فوسفوديستريس أنا (NPP1)، جنبا إلى جنب مع أنكيرين (عنك)، يتحكم تركيز بيروفوسفات غير العضوية (PPأنا) 6-PPأنا، مثبط قوية لتشكيل هكتار، وهي تحلل قبل تناب؛ NPP1 هيدروليزيس تريفوسفاتيس النوكليوتيدات لتشكيل PPأنا بينما الصادرات PPأنا عنك الخلية إلى إدارة المحتوى في المؤسسة. قد تنظم نسبة Pi/PPi الاباتيت تشكيل7،8 مع9من النتائج المرضية المحتملة.
هو إثراء الغشاء أم في أيون النقل البروتينات التي تيسر الترسيب الأولية من الكالسيوم والفوسفات داخل MVs أثناء عملية نويات (الشكل 1). نقل الفوسفات 1 (PiT) يساعد على إدماج فأنا ولدت في الفضاء بيريفيسيكولار إلى10،MVs11. يمكن إشراك أنيكسينس في الربط والنقل من Ca2 + وفي العملية الفيزيائية الحيوية التي يبدأ تمعدن في12،التجويف المتوسط13. أننا نفضل الفرضية، اقترح في وقت سابق التمعدن داخل الحويصلات إينتراسيتوبلاسميك من نويات الداخلية من اباتيت داخل MV قبل انتشاره في14،ECM15. وأكد النمذجة في المختبر تحريض Ca2 +/Pأنا مجمعات تشكيل في بروتيوليبوسوميس مصنوعة من PS و AnxA516. قد يشير هذا إلى أن التراكم من Ca2 +, فأنا، تمتلك مجمعات AnxA5 و PS في اطواف الدهن من مثل زغيبات ميمبرانيسريبريسينت لب نويات (NC) الاباتيت داخل ممقابل أنيكسينس وطنب أيضا ملزمة الكولاجين القدرات التي قد تكون مفيدة في وضع MVs على طول ألياف الكولاجين، وتنشيط الدعوة التمعدن في إدارة المحتوى في المؤسسة. أ فتوين وأوستيوبونتين (أوبن)17, معروفة كمثبطات لتشكيل الاباتيت التي قد تبطئ انتشار التمعدن على سقالة collagenous. التنو ونشر الأحداث متميزة، والسابق السابق لهذا الأخير، وكلاهما قد تكون ذات صلة بعملية تمعدن المرضية.
لاكتشاف كيف يمكن تغيير كيمياء مجمعات فوسفات الكالسيوم التمعدن الفسيولوجية وتكلس حمل خارج الرحم، من الضروري تحديد المعادن التي تنتجها الخلايا. أباتيتيس مجموعة من الكالسيوم والفوسفات التي تحتوي على معادن مع كريستال العام وحدة خلية الصيغة Ca10(ص4)6X2، حيث X = Cl، واو، أوهايو. كانت تصنف على النحو التالي18: فلوراباتيتي (FA) Ca10(ص4)6و2،10(ص4) تشلوراباتيتي (CA) Ca6Cl2 وهيدروكسيباتيت (ها) Ca10(ص. ب4 )6(OH)2.
اختيار خطوط الخلايا أوستيوبلاست للحث على تشكيل المعادن حاسم، حيث يسلك كل خط خلية مكانة متميزة التمعدن. في هذا التقرير، قارنا التنو المعادن بنموذجين الخلية البشرية المحددة التمعدن: هفوب أوستيوبلاستيك 1.19 الخلايا والخلايا osteosarcoma ساوس-2. Osteosarcoma المستمدة من الخلايا تستخدم عادة كنماذج أوستيوبلاستيك وخلايا سوس-2 حافظوا على الطابع أوستيوبلاستيك الأكثر نضجاً19 بينما خلايا غير متمايزة هفوب الأجنة البشرية تستخدم على نطاق واسع كنموذج لطبيعية أوستيوبلاستيك 20من التفريق. حللت ملفاتهم التمعدن بأساليب مختلفة: تلطيخ الأليزارين الأحمر-S (ع-ق) والأشعة فوق البنفسجية التصور الضوء (الأشعة فوق البنفسجية) وانتقال تصوير الميكروسكوب الإلكتروني (TEM)، الأشعة السينية المشتتة الطاقة سليكية (EDX) كوانتيتيشن وأيون رسم الخرائط. ميزة ال EDX عبر التقنيات البديلة المستخدمة في الدراسات السابقة أنه يعطي النتائج الكمية والنوعية لاستبدال أيون في الاباتيت بلورات4،5،21. وكان الهدف الشامل المتمثل في استخدام ال EDX للعثور على طريقة بسيطة للتصوير والتحديد الكمي لتوزيع أيونات Ca، و و Cl في المعادن المختلفة من أنواع مختلفة من الخلايا خلال مراحل متميزة من عملية تعدين. هذا الأسلوب قد استخدمت بنجاح، على سبيل المثال، لرصد تفاعل جسيمات نانوية الزنك مع المواد الكيميائية تتعايش وآثارها مجتمعة على الكائنات المائية22. وفي دراسة أخرى، اتسمت فوتوكاتاليست نحاس في مواد التيتانيوم في محلول مائي على نطاق واسع عن طريق قياس الطيف الكتلي بلازما الحث يقترن الانبعاثات الضوئية (برنامج المقارنات الدولية-OES)، فيسيسوربشن N2 (الرهان)، زرد، تجاه الأشعة فوق البنفسجية DRS، قدم الأشعة تحت الحمراء، رامان التحليل الطيفي، وال EDX والفوتوكهروكيميائيه قياسات23. وكان هدفنا المقارنة بين الأصل وخصائص من حويصلات والمعادن في اثنين من خطوط الخلية لفهم الآلية التي يتحكم التمعدن خلال التمايز العظمى.
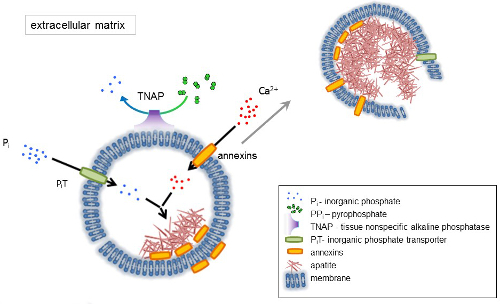
الشكل 1 . مخطط للخطوات الأولية التمعدن في الخلايا العظمى التي تنطوي على تركيب البروتينات المصفوفة خارج الخلية (ECM) والإفراج عن مصفوفة حويصلات (MVs) من الغشاء. MVs تتراكم الكالسيوم من خلال عمل الكالسيوم ملزم البروتينات وأننيكسينس والفوسفات، عن طريق العمل نقل الفوسفات غير العضوي (PiT) متبوعاً بنشاط الأنسجة غير محددة القلوية الفوسفاتيز (تناب)، الذي ديفوسفوريلاتيس PPأنا إلى فأنا، مما ييسر التنو الاباتيت. ثم MVs تتفكك والإفراج عن أباتيتيس إلى الوسط خارج الخلية. هو استمرار التمعدن بإمدادات ثابتة من فأنا و Ca2 + في المتوسط خارج الخلية4،5. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
