El hueso es un tipo de tejido conectivo compuesto de dos partes: orgánica (células y fibras de colágeno) y minerales (compuestos de calcio y fosfato). Los principales componentes minerales en los huesos son apatitas1. Diferentes tipos de células competentes de mineralización en el hueso (osteoblastos), dientes (odontoblastos) y cartílago (condrocitos) regulan los pasos iniciales de mineralización produciendo proteínas de la matriz extracelular (ECM) y liberación de matriz vesículas (MVs) (figura 1). MVs son vesículas de 100 a 300 nm de diámetro que acumulan calcio y fosfato facilita la nucleación de apatita y posteriormente se unen al colágeno2,3. Entonces, MVs se desintegran para liberar apatitas al medio extracelular. Las apatitas continúan creciendo en contacto con las fibras de colágeno y forman la matriz ósea. La mineralización es sostenida por el suministro constante de P y Ca2 + en el medio extracelular. Algunos datos publicados recientemente apoyan nuestro modelo4,5. Los tejidos blandos no mineralizarse bajo condiciones fisiológicas. Sin embargo, la calcificación ectópica puede ocurrir en condiciones patológicas tales como calcificación vascular3. Las células vasculares que adquieren el fenotipo de osteoblastos pueden producir MVs que inducen la nucleación de apatitas e inician la mineralización en las capas íntimas y mediales de la pared de los vasos sanguíneos. Desde calcificación ectópica se asemejan a endocondral normal mineralización3, entender los mecanismos moleculares de la mineralización de las células óseas y condrocitos deben proporcionar algunas pistas sobre la calcificación ectópica de los tejidos blandos que son formado.
El desarrollo de tejidos esqueléticos está regulado por diversas enzimas, factores de crecimiento y promotores o inhibidores de la mineralización. La acción antagónica de la fosfatasa alcalina no específica de tejido (TNAP) (figura 1) y ectonucleotide pirofosfatasa/fosfodiesterasa I (NPP1), junto con ankyrin (ANK), controla la concentración de pirofosfato inorgánico (PP) 6. PP, un potente inhibidor de la formación de HA, es hidrolizado por TNAP; NPP1 hidroliza trifosfatos de nucleótidos para formar PP mientras ANK las exportaciones de PP de la célula al ECM. La proporción Pi/PPi puede regular la formación de apatita7,8 con posibles consecuencias patológicas9.
La membrana de MV se enriquece en proteínas de transporte de iones que facilitan la precipitación inicial de calcio y fosfato en el MVs durante el proceso de nucleación (figura 1). El transportador de fosfato 1 (hoyo) ayuda a incorporar P genera en el espacio perivesicular en la MVs10,11. Anexinas pueden estar implicado en el atascamiento y el transporte de Ca2 + y en el proceso biofísico que inicia la mineralización en el MV lumen12,13. Privilegiamos la hipótesis, sugerida anteriormente, para la mineralización dentro de vesículas intracitoplasmáticas de nucleación interna de apatita dentro de la MV antes de su propagación en el ECM14,15. Modelado in vitro confirmaron la inducción de la Ca2 +/P complejos formación en proteoliposomes de PS y AnxA516. Esto puede indicar que la acumulación de Ca2 +, P, complejos AnxA5 y PS en las balsas lipídicas de las microvellosidades-como membranesrepresent el núcleo nucleación (NC) de apatita dentro de Mvs anexinas y TNAP también poseen colágeno vinculante capacidades que pueden ayudar en la colocación de MVs a lo largo de las fibras de colágeno y en estimular la propagación de la mineralización en el ECM. Fetuin A y17de la osteopontina (OPN), son conocidos como inhibidores de la formación de apatita que puede ralentizar la propagación de la mineralización en el andamio colagenosa. Nucleación y propagación son eventos distintos, el primero anterior a éste, y ambos pueden ser relevantes para el proceso de mineralización patológica.
Para descubrir cómo puede cambiar la química de complejos de fosfato de calcio calcificación ectópica y mineralización fisiológica, es necesario identificar los minerales producidos por las células. Apatitas son un grupo de calcio y fosfato que contiene minerales con el cristal general unidad celular fórmula Ca10(PO4)6X2, donde X = Cl, F, OH. Se clasifican de la siguiente manera18: Fluorapatito (FA) Ca10(PO4)6F2y chlorapatite (CA) Ca10(PO4)6Cl2 de hidroxiapatita (HA) Ca10(PO4 )6(OH)2.
La elección de las líneas celulares de osteoblastos para inducir la formación de minerales es crucial, puesto que cada línea celular exhibe un perfil distinto de mineralización. En este informe, en comparación con la nucleación de minerales por dos modelos de células humanas las de mineralización: células osteoblásticas hFOB 1.19 y células de osteosarcoma Saos-2. Las células derivadas de osteosarcoma se utilizan comúnmente como modelos osteoblásticos y células Saos-2 conservan la más madura de carácter osteoblástico19 mientras que las células indiferenciadas hFOB fetal humano son ampliamente utilizadas como modelo para osteoblástica normal diferenciación20. Sus perfiles de mineralización se analizaron mediante diferentes métodos: tinción rojo de alizarina-S (AR-S), ULTRAVIOLETA (UV) luz la visualización, la proyección de imagen de microscopía electrónica (TEM) transmisión, cuantificación de microanálisis (EDX) de rayos x dispersivo energía y ion asignación. La ventaja de TEM-EDX sobre alternativas técnicas utilizadas en estudios previos es que da resultados cuantitativos y cualitativos de la sustitución de iones en cristales de apatita4,5,21. El objetivo de utilizar las TEM-EDX era encontrar un método simple para la proyección de imagen y cuantificación de la distribución de los iones Ca, F y Cl en varios minerales de diferentes tipos de células en distintas etapas del proceso de mineralización. Este método ha sido utilizado con éxito, por ejemplo, para monitorear la interacción de las nanopartículas de zinc con productos químicos coexistentes y sus combinados efectos en organismos acuáticos22. En otro estudio, un fotocatalizador cobre en materiales de titanio en solución acuosa se caracterizó ampliamente por medio de espectrometría de emisión óptica de plasma acoplado inductivamente (ICP-OES), N2 physisorption (BET), DRX, DRS de UV-vis, FT-IR, Raman Espectroscopia, TEM-EDX y fotoelectroquímicos medidas23. Nuestro objetivo fue comparar el origen y las propiedades de las vesículas y los minerales en dos líneas celulares para entender el mecanismo que controla la mineralización durante la diferenciación ósea.
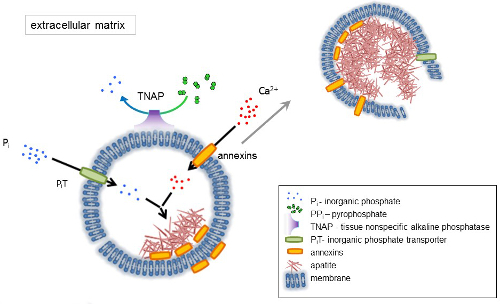
Figura 1 . Esquema de los pasos iniciales de la mineralización en las células óseas que implican la síntesis de proteínas de matriz extracelular (ECM) y liberación de vesículas de la matriz (MVs) de la membrana. MVs acumulan calcio a través de la acción de proteínas calcio, anexinas y fosfato, a través de la acción de un transportador de fosfato inorgánico (PiT) seguido por la actividad de tejido no-fosfatasa alcalina específica (TNAP), que desfosforila PP a P, facilitando así el nucleation de la apatita. Entonces, MVs se desintegren y liberen apatitas al medio extracelular. La mineralización es sostenida por el suministro constante de P y Ca2 + en el medio extracelular4,5. Haga clic aquí para ver una versión más grande de esta figura.
