Oligodendrocytes (OLs) sont les cellules myélinisantes du système nerveux central (SNC)1. L’isolement et la culture des oligodendrocytes primaires dans un environnement fortement réglementé est un outil précieux pour l’étude in vitro de l’élaboration des oligodendrocytes ainsi que la biologie des maladies telles que la sclérose2 démyélinisantes . Cela nécessite un oligodendrocyte efficace et robuste d’isolement et de la culture la méthode3. Dans cette étude, nous avons profité de l’expression d’un marqueur de surface de cellule oligodendrocyte distinctif pour mettre en œuvre une technique d’isolation modifiée qui est rapide et précis.
Quatre étapes distinctes de la maturation des oligodendrocytes ont été identifiés, chacun caractérisé par l’expression des marqueurs de surface de cellules distinctes pour chaque stade de développement (Figure 1). Ces marqueurs de surface cellulaire peuvent être reconnus par des anticorps spécifiques4,5et peuvent être utilisés pour isoler les langues officielles à des étapes spécifiques. Dans un premier temps, cellules de précurseurs d’oligodendrocytes (OPCs) ont la capacité à proliférer, de migrer et de façon explicite de facteur de croissance dérivé des plaquettes (PDGF-Rα) de récepteur6, ganglioside A2B5, protéoglycane NG27,8 , polysialic acide-neural cell adhesion molécule9 et gras-acid-binding protein 7 (FABP7)10. OPCs ont une morphologie bipolaire avec peu de processus abrégé émanant des pôles opposés du corps cellulaire, qui est caractéristique des précurseurs neuronaux de cellules11.
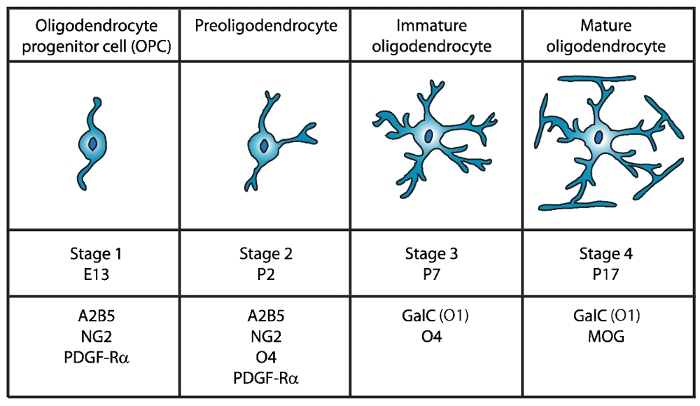
Figure 1 : Expression de marqueurs de surface cellulaire lors du développement d’oligodendrocyte souris. Opération survie cellulaire marqueurs de surface tels que A2B5, GalC (O1), NG2, O4 et PDGF-Rα peuvent servir à isoler spécifiquement les oligodendrocytes au stade de développement spécifique en utilisant des anticorps spécifiques. Veuillez cliquer ici pour visionner une version agrandie de cette figure.
Dans un deuxième temps, OPCs donnent lieu à la preoligodendrocytes et exprimer à la membrane cellulaire non seulement des marqueurs OPC, mais aussi le Sulfoglycosphingolipides (un galactolipide sulfatée) reconnu par le O4 anticorps12,13et la protéine de GPR1714, qui persiste jusqu’au stade d’oligodendrocytes immatures (OL). À ce stade, preoligodendrocytes étendre les processus abrégés multipolaires. Preoligodendrocytes sont l’étape majeure d’OL à jour après la naissance 2 (P2) dans la substance blanche cérébrale de rat et de souris avec une population mineure d’immatures OLs15.
Au cours de la troisième phase, OLs immatures continuent à exprimer O4, perdre l’expression des marqueurs A2B5 et NG2 et commencent à exprimer les galactocérébrosides C16. À ce stade, opération Survie au Soudan se sont engagés à la lignée oligodendrocyte et deviennent des cellules post-mitotiques avec des branches bien ramifiées17,18. À cette époque les premières cellules MBP+ on observe15,19,20,21et OL immature constituent plus de 80 % de la matière blanche rongeur à P7. Par conséquent, isolement des langues officielles à P7 pourrait garantir cellulaire à haut rendement.
Dans la finale et quatrième étape du développement de l’OL, OLs matures expriment myélinisantes protéines (la protéine basique de la myéline (MBP), protéine protéolipidique (PLP), glycoprotéine myéline associée (MAG) et la myéline oligodendrocyte glycoprotéine (MOG)22,23 ,24,25,26. À ce stade, OLs matures s’étendent les membranes ce compact de forme Envellopants gaines autour des axones et peuvent myéliniser. Cela coïncide avec l’observation que, dans le cerveau de rat et de souris, les cellules MBP+ deviennent plus en plus abondantes à P1419,20,21.
Depuis le premier isolement des oligodendrocytes par fifi et ses collègues en 196727, plusieurs méthodes pour l’isolement des langues officielles de la CNS du rongeur ont été appliquées, y compris immunopanning28,29,30, cellule activée par fluorescence triant (FACS) exploitant cell surface spécifique antigènes28,31, centrifugation en gradient différentiel32,33,34,35 et secouant méthode fondée sur l’adhésion différentielle des différents CNS glia36,37. Cependant, les méthodes existantes de la culture ont des limites, notamment en termes de pureté, de rendement et de temps requis pour exécuter les procédures38. Par conséquent, des méthodes d’isolation plus efficaces pour les oligodendrocytes sont nécessaires.
Dans cet article, nous présentons un simple et méthode de sélection efficace pour l’isolation immunomagnetic de stade trois O4+ cellules preoligodendrocytes de petits souris néonatales. Cette méthode est une modification des techniques signalées par Emery et al. 39 et Dincman et al. 40 et assure une oligodendrocyte préparation une pureté supérieure à 80 % en environ 4 h.
