Gli organismi fotosintetici come le diatomee devono affrontare condizioni di luce mutevoli e rispondere con meccanismi sofisticati di acclimatazione che sostengono ad alta efficienza fotosintetica e proteggono da danno foto-ossidativo causato dalla luce eccessiva. Un importante processo di luce-protettivo negli eucarioti fotosintetici è l’alta energia tempra (qE) di luce assorbita che si presenta come il principale contributo per la tempra non fotochimica (NPQ) sotto condizioni di luce lo stress1,2 ,3. Il luce complessi antenna raccolta (LHC) sono coinvolti nella regolazione delle vie di trasferimento di energia di eccitazione. In risposta a luce alta indotta basso pH nel lume cloroplasto, gli interruttori del sistema antenna dalla luce raccolta allo stato dissetante. Questo stato che tende a dissipare energia protegge fotosistema (PS) e altri complessi nella membrana tilacoide da foto-ossidazione. Negli eucarioti fotosintetici, la qE solitamente è indotta da due fattori1,2,3. Un fattore è la luce specializzata raccolta della proteina che risponde al basso pH. La proteina PsbS induce il qE in più piante4. LhcSRs5, modulata da attività PsbS, indurre il qE alghe verdi6. Le diatomee possiedono proteine Lhcx-like che strutturalmente correlata alla LHCSRs7,8,9,10.
Il secondo fattore di qE è il ciclo della xantofilla dove i carotenoidi dell’antenna sono convertiti in un formato di foto-protettivo dal de-epossidazione e ripristinati da epossidazione. Nelle piante e alghe verdi, violaxantina viene convertito in zeaxantina. A diatomee, diadinoxanthin viene convertito in diatoxanthin, che quindi correla con il grado di NPQ11. La luce della diatomea raccolta antenna possiede alcune peculiarità, anche se è evolutiva legate alla pianta e LHCs d’alghe. L’interruttore da luce raccolta foto-protezione è estremamente veloce e la capacità NPQ è superiore rispetto a piante12. Questo potrebbe essere una ragione perché le diatomee sono molto successo in diverse nicchie ecologiche, in modo che essi sono responsabili fino al 45% della produzione primaria netta oceanico13. Di conseguenza, della diatomea luce sistemi di raccolta sono un interessante oggetto di ricerca di fotosintesi.
Diatomee, come le specie centric Cyclotella meneghiniana, possiedono luce intrinseca thylakoid prende i pigmenti hanno sistemi di raccolta associare – fucoxantina, clorofilla (chl) a e c, quindi FCP. luce raccolta proteine, quali FCPs, sono incorporato nel sistema della membrana tilacoide formati da diversi strati di membrana. Diatomee formano le fasce dei tre tilacoidi. Questo complesso situazione rende difficile per loro studiare a livello molecolare nella membrana tilacoide. Inoltre, molti componenti contribuiscono alla regolazione della luce raccolta (Vedi sopra). Di conseguenza, in molti approcci, i complessi sono stati isolati dalla membrana utilizzando un detergente delicato, come n-dodecil-β-D-maltopyranoside (β-DDM), che solubilizzano la membrana, ma mantenere intatti i complessi FCP. Molti studi spettroscopici sono stati eseguiti utilizzando solubilizzate FCP per indagare intramolecolare energia trasferimento14,15,16,17. Tuttavia, questo approccio ex era limitato poiché il regolamento del trasferimento di energia ha bisogno eccitoniche interazione con altri complessi antenna o fotosistema. Quindi, questi generi di studi non possono essere effettuati con solubilizzate complessi perché l’interazione fra i complessi è perso.
Una caratteristica importante nella regolazione dell’antenna è il “affollamento molecolare” del fotosistema nella membrana tilacoide18e dell’antenna. Precedentemente, è stato realizzato un approccio semplice per simulare questo effetto in vitro. Il detersivo è stato rimosso, che porta all’aggregazione casuale di complessi antenna. Anche se alcuni dati ragionevoli è stati ottenuti da questo approccio17,19, la rimozione del detersivo non riflette la situazione in vivo e presenta alcune limitazioni, poiché i complessi non interagiscono nel loro terziari regolari struttura.
L’uso dei liposomi supera molte delle limitazioni ex. La struttura terziaria è ancora completamente intatta. La membrana del liposoma fornisce un ambiente quasi nativo per i complessi dell’antenna. La membrana separa l’interno del liposoma dall’ambiente esterno. Con questi mezzi, liposomi forniscono due scomparti di reazione per studi di gradienti ionici e pH pure per quanto riguarda i processi di trasporto. Ulteriormente, i parametri del sistema sperimentale possono essere controllati più facilmente per gli studi nella membrana tilacoide. Liposomi già sono stati indicati per essere un ottimo strumento per lo studio di complessi fotosintetici. Degli obiettivi principali in passato era sulla pianta LHC dove l’effetto della composizione lipidica alterato è stato testato su LHC II20. In altri approcci, interazione proteina-proteina tra diversi LHC II sono stati studiati21. Inoltre, alcuni studi in alghe verdi sono stati effettuati che descrivono il clustering spontanea tra LHC22. Considerando l’importanza di diatomee per gli ecosistemi acquatici, relativamente pochi studi sono stati effettuati con complessi antenna di diatomee. Due studi hanno studiato i complessi antenna della centrica Cyclotella meneghiniana, dove il clustering della FCP antenna23 e reattività di FCP a gradienti elettrochimici24 sono stati indicati. Così, i liposomi sono un ottimo strumento per lo studio della diatomea antenne e loro interazione e regolazione in condizioni quasi native. I liposomi sono versatili da molte condizioni come composizione lipidica, liposoma dimensioni, densità di proteina e la fase acquosa circostante può essere controllata. Inoltre, il metodo richiede basse quantità di campioni. Il sistema sperimentale è robusto e altamente riproducibili. La compartimentazione dei liposomi permette lo studio di pH e gradienti ionici, che sono importanti fattori nella regolazione dei complessi antenna.
Qui, descriviamo l’isolamento di complessi antenna FCP da c. meneghiniana e la loro incorporazione in liposomi con composizione lipidica naturale thylakoid. Inoltre, forniamo dati esemplari per la caratterizzazione spettroscopica di FCP solubilizzate e confrontarli con FCP in liposomi. Il metodo riassume conoscenze e protocolli standardizzati ottenuti dai miglioramenti di Gundermann e Büchel 201223, Natali et al 201622e Ahmad e Dietzel 201724.
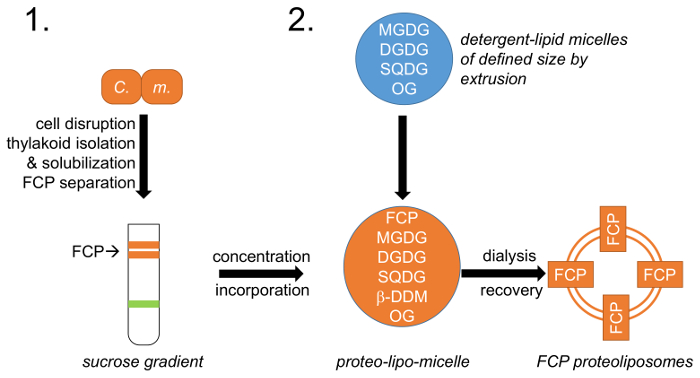
Figura 1: rappresentazione schematica del flusso di lavoro. (1) fa riferimento al paragrafo 1 che descrive la crescita delle cellule, rottura e tilacoidale isolamento con seguito di FCP separazione su gradienti di densità di saccarosio; M. c. –Cyclotella meneghiniana cellule. (2) preparazione della miscela di lipidi naturali thylakoid (MGDG, DGDG e SQDG) descritto nel paragrafo 2 e creazione di micelle lipidiche-detergente con octylglycoside (OG). Una dimensione definita del lipido-micella avviene mediante estrusione utilizzando membrane di un diametro dei pori definiti. FCP e lipido-micelle sono unificate presso un lipide predefinito: rapporto proteine e i detergenti OG e β-DDM vengono rimossi tramite controllata dialisi formando FCP proteoliposomi. Clicca qui per visualizzare una versione più grande di questa figura.
