Veel biologische processen zich voordoen op de sites van cel-cel interacties, bijvoorbeeld cel-cel adhesie1,2,3, cel-cel fusion4 en cellulaire erkenning5. Dergelijke gebeurtenissen zijn vooral belangrijk bij de ontwikkeling van meercellige organismen en voor cel-cel communicatie, bijvoorbeeld tijdens de immuunrespons. Deze processen zijn meestal gemedieerd door eiwitten die zijn gelokaliseerd aan het oppervlak, dat wil zeggen, op het plasma-membraan (PM) van naburige cellen en specifieke interacties op de cel-cel-contactpersoon die precies geregeld in ruimte en tijd te ondergaan. In veel gevallen deze interacties zijn directe homo- of heterotypic eiwit-eiwitinteractie trans interacties, maar mogelijk ook ionen of liganden hoedanigheid van extracellulaire linkers1. Hoewel het van fundamenteel belang, is er een gebrek assays indringende deze specifieke eiwit-eiwitinteractie rechtstreeks in de eigen omgeving van levende cellen. Veel methoden vereisen of verstoring van de cel (bijvoorbeeld biochemische tests zoals co-immunoprecipitation6), fixatie (bijvoorbeeld aantal super resolutie optische microscopie technieken en elektronenmicroscopie van cel-cel contact op met7), of zijn niet-specifieke, bijvoorbeeld aggregatie / hechting testen8,9. Om dit probleem te overwinnen, zijn fluorescentie technieken doorgevoerd op basis van fluorescentie resonance energy transfer (FRET)10 of fluorescentie complementatie11. Deze methoden vereisen echter om voldoende kleine afstanden tussen fluorophores, fluorescerende etiketten aan de extracellulaire kant van de eiwitten10, mogelijk verstoren trans interacties.
Hier presenteren we een alternatieve fluorescentie gebaseerde assay voor eiwit-eiwitinteractie op cel contacten. Deze aanpak combineert fluorescentie cross-correlatie benaderingen (scannen fluorescentie cross-correlatie spectroscopie (sFCCS), cross-correlatie nummer en helderheid (ccN & B)) en het mengen van cellen uiten van een constructie van de fusie voor het eiwit van belang, bijvoorbeeld een hechting-receptor. De onderzochte receptoren in de twee interagerende cellen worden aangeduid met twee spectraal gescheiden fluorescente proteïnen (FPs), van de intracellulaire kant (Zie figuur 1A).
De werknemer methoden zijn gebaseerd op de statistische analyse van de fluorescentie schommelingen veroorzaakt door de diffusive motie van fluorescerende fusion eiwitten door het focal volume van een confocale laser scanning microscoop. Meer in detail sondes de bepaling de co verspreiding van de proteïnen van belang in beide naburige PMs op cel contacten. Als de eiwitten ondergaan trans interacties, zal deze trans -complexen fluorescente proteïnen uitstoten in beide spectrale kanalen, waardoor gecorreleerde fluorescentie schommelingen van beide vervuilers dragen. Aan de andere kant, als geen binding treedt op, zal het aantal schommelingen van eiwitten bij het omgaan met PMs onafhankelijk zijn, waardoor geen gecorreleerde schommelingen. De overname kan op twee manieren worden uitgevoerd: 1) sFCCS is gebaseerd op een lijn-vormige scan door de contactpersoon van de cel-cel en effectief onderzoekt de interacties in een plek gelegen in de regio contact. Door middel van een temporele analyse van fluorescentie schommelingen biedt sFCCS ook dynamica informatie, dat wil zeggen, de coëfficiënten van de diffusie van eiwitcomplexen; 2) ccN & B is gebaseerd op een pixel-wise analyse van een reeks beelden die zijn verworven op de cel contact regio’s. Het heeft de mogelijkheid om te sonderen en kaart interacties langs de hele Neem contact op met de regio (in één brandvlak), maar levert geen informatie over dynamiek. Beide methoden kunnen worden gecombineerd met een analyse van de moleculaire helderheid, dat wil zeggen, de gemiddelde fluorescentie-signaal in de tijdseenheid uitgestoten door één verspreiden van eiwitcomplexen en schattingen van de stoichiometrie van eiwitcomplexen op dus op te leveren de contactpersonen van de cel.
In dit artikel geven wij gedetailleerde protocollen voor monstervoorbereiding, kalibreren van het instrument, data-acquisitie en analyse uit te voeren van de voorgestelde bepaling over een commerciële confocale laser scanning microscoop. De experimenten kunnen worden uitgevoerd op elk instrument voorzien van foton tellen of analoge detectoren en een doelstelling met hoge numerieke diafragma. We verder bespreken kritische stappen van het protocol en correctie regelingen voorzien in verschillende processen veroorzaken artefactual signaal schommelingen, bijvoorbeeld detector geluidshinder, photobleaching of cel verkeer. Oorspronkelijk ontwikkeld om sonde interacties tussen Adherente cellen, de bepaling voor schorsing cellen kunnen worden gewijzigd of aangepast aan membraan modelsystemen, bijvoorbeeld gigantische unilamellar blaasjes (GUVs) of gigantische plasma membraan blaasjes (GPMVs), waardoor de kwantificering van interacties in verschillende lipide omgevingen of bij gebreke van een georganiseerde cytoskelet12,13.
Het scannen van fluorescentie cross-correlatie spectroscopie is een gewijzigde versie van fluorescentie cross-correlatie spectroscopie14 en werd specifiek ontworpen om de sonde langzaam diffusive dynamiek in lipide membranen15. Het is gebaseerd op een lijn-scan overname loodrecht op de PM met de fluorescente proteïnen van belang. Om de sonde twee anders gelabelde eiwit soorten interacties, wordt de verwerving uitgevoerd in twee spectrale kanalen met behulp van twee laserlijnen en twee detectie Vensters voor spectraal gescheiden fluorophores. Vanwege de dynamiek van de trage verspreiding van eiwitten in de PM (D≤ ~ 1 µm2/s), een cross-talk-gratis meting kan worden uitgevoerd door afwisselend de regeling van de excitatie van lijn tot lijn15. De analyse begint met: 1) een uitlijning algoritme corrigeren voor laterale cel verkeer op basis van block-wise gemiddeld van ~ 1000 lijnen, 2) bepaling van de positie met maximale fluorescentie signaaldat wil zeggen, de PM positie, in elk blok en 3) verschuiven van alle blokken te een gemeenschappelijke oorsprong12,15, afzonderlijk in elk kanaal. Vervolgens wordt een automatische selectie van pixels die overeenkomen met de PM uitgevoerd door het selecteren van de regio in het midden van een Gaussiaanse pasvorm van de som van alle uitgelijnde lijnen (d.w.z., centrum ± 2.5σ). Integratie van het signaal in elke regel levert de membraan fluorescentie tijdreeksen F(t) in elk kanaal (g = groen kanaal, r = rode kanaal). Opmerking dat de pixelgrootte moet klein genoeg, bijvoorbeeld < 200 nm, te reconstrueren van de vorm van het punt verspreiden op functie en vindt haar center, overeenkomt met de positie van de PM. In de aanwezigheid van aanzienlijke photobleaching, de fluorescentie tijdreeksen in elk kanaal kan worden gemodelleerd met een dubbel-exponentiële functie en vervolgens gecorrigeerd met behulp van de volgende formule:16
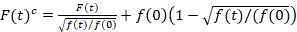 . (1)
. (1)
Het is belangrijk op te merken dat deze formule effectief corrigeert zowel de amplitudes en de diffusie tijden verkregen correlatie analyse van F(t)c, vergeleken met de ramingen van de parameter die uit de niet-gecorrigeerde F(t)zou worden verkregen. Vervolgens, de auto – en cross-correlatie functies (ACFs / CCFs) van de fluorescentie signalen worden berekend:
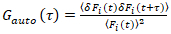 , (2).
, (2).
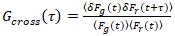 , (3).
, (3).
waar δFik = Fik(t) – 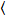 Fik(t)
Fik(t)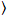 en ik = g, r.
en ik = g, r.
Het model van een twee-dimensionale diffusie is dan voorzien tot alle functies van de correlatie (CFs):
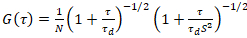 . (4)
. (4)
Hier, geeft N het aantal fluorescente proteïnen in de omvang van de observatie en τd de diffusie-tijd voor elk kanaal. Dit model houdt rekening dat in de beschreven experimentele setting, verspreiding van eiwitten in de PM treedt op in het x-z vlak, in tegenstelling tot de veel gebruikte configuratie voor fluorescentie correlatie spectroscopie (FCS) experimenten op membranen sonderen verspreiding in de x-y-vlak van de confocal volume17. De taille w0 en de structuur factor S, met een beschrijving van de rek wz van het focal volume in z, S = wz/w0, een punt FCS kalibratie meting uitgevoerd met spectraal soortgelijke kleurstoffen en dezelfde optische instellingen worden verkregen met behulp van reeds beschikbare waarden voor de diffusie-coëfficiënt Dkleurstof:
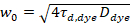 , (5).
, (5).
waar τd, kleurstof de tijd gemeten gemiddelde verspreiding van de kleurstof moleculen is, verkregen uit de montage van een model voor driedimensionale verspreiding op gegevens, rekening houdend rekening overgangen van een breuk T van alle N -moleculen aan een triplet staat met een tijdconstante ττ:
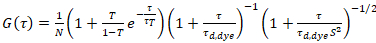 . (6)
. (6)
Tot slot zijn diffusie coëfficiënten (D), moleculaire helderheidswaarden (ε) en de relatieve cross-correlatie van sFCCS gegevens (rel.cc.) als volgt berekend:
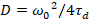 , (7).
, (7).
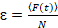 , (8).
, (8).
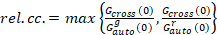 , (9).
, (9).
waar GKruis(0) is de amplitude van de functie cross-correlatie en 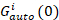 is de amplitude van de autocorrelatiefunctie in het ik-th kanaal.
is de amplitude van de autocorrelatiefunctie in het ik-th kanaal.
Deze definitie van de relatieve cross-correlatie, dat wil zeggen met behulp van max in plaats van betekenen in vergelijking 9, houdt rekening dat het maximum aantal complexen van twee eiwit soorten waarvan de aanwezigheid in verschillende concentraties wordt beperkt door de soorten waarvan de aanwezigheid in een lager nummer.
Cross-correlatie nummer en helderheid is gebaseerd op een analyse van het moment van de intensiteit van de fluorescentie voor elke pixel van een afbeeldingsstapel verworven na verloop van tijd op een vaste positie in de steekproef, meestal bestaande uit ~ 100-200 frames, met twee spectrale kanalen () g = groen kanaal, r = rode kanaal). Ten opzichte van de temporele gemiddelde ikik en variantie 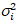 , de moleculaire helderheid εi en nummer nik worden berekend in elke pixel en spectrale kanaal (Ik = g, r)18:
, de moleculaire helderheid εi en nummer nik worden berekend in elke pixel en spectrale kanaal (Ik = g, r)18:
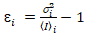 , (10).
, (10).
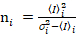 . (11)
. (11)
Het is belangrijk op te merken dat de gegeven vergelijkingen voor de ideale geval voor een ware foton-tellen detector gelden. Voor analoge detectiesystemen gelden de volgende vergelijkingen19,20:
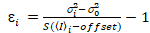 , (12).
, (12).
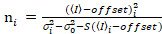 . (13)
. (13)
Hier, S is de omrekeningsfactor tussen gedetecteerde fotonen en de opgenomen digitale graven, 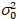 is de uitlezing lawaai en offset verwijst naar de verschuiving van de intensiteit van de detector. In het algemeen, deze hoeveelheden moeten worden gekalibreerd, voor elk type van de detector, gebaseerd op de meting van de variatie van de detector als een functie van intensiteit voor constante verlichting19, bijvoorbeeld een reflecterend oppervlak van metalen of gedroogde kleurstof oplossing. De verschuiving kan worden bepaald door het meten van de graaf-tarief voor een monster zonder excitatie licht. Door het uitvoeren van een lineaire regressie van de afwijking van de detector-geassocieerde
is de uitlezing lawaai en offset verwijst naar de verschuiving van de intensiteit van de detector. In het algemeen, deze hoeveelheden moeten worden gekalibreerd, voor elk type van de detector, gebaseerd op de meting van de variatie van de detector als een functie van intensiteit voor constante verlichting19, bijvoorbeeld een reflecterend oppervlak van metalen of gedroogde kleurstof oplossing. De verschuiving kan worden bepaald door het meten van de graaf-tarief voor een monster zonder excitatie licht. Door het uitvoeren van een lineaire regressie van de afwijking van de detector-geassocieerde 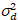 versus intensiteit (ik) perceel, S en kan worden bepaald19:
versus intensiteit (ik) perceel, S en kan worden bepaald19:
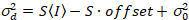 . (14)
. (14)
Tot slot, de cross-correlatie helderheid in elke pixel wordt berekend en wordt gedefinieerd in het algemeen als21
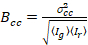 , (15).
, (15).
waar 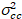 is de Kruis-variantie
is de Kruis-variantie 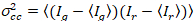 .
.
Om het filteren van langlevende schommelingen, worden alle ccN & B berekeningen uitgevoerd na een boxcar filtering, onafhankelijk voor elke pixel22. Kort, ni, εik (Ik = g, r) en Bcc in glijden segmenten van bijvoorbeeld 8-15 frames worden berekend. De aldus verkregen waarden kunnen vervolgens om te verkrijgen van de laatste pixel aantal en helderheid waarden worden gemiddeld.
Stoichiometrie analyse
Om te kunnen inschatten van de stoichiometrie van eiwitcomplexen op cel contactpersonen, kan de moleculaire helderheid afzonderlijk worden geanalyseerd in elke spectrale kanaal voor de sFCCS of ccN & B gegevens. In sFCCS, wordt een helderheidswaarde verkregen per meting in elk kanaal. In ccN & B, een histogram van de helderheid van alle pixels die overeenkomen met de contactpersoon van de cel wordt verkregen en de waarde van de gemiddelde (of mediaan) kan worden gebruikt als vertegenwoordiger helderheid voor de meting. Door het uitvoeren van de dezelfde analyse op een monomeer verwijzing, kunnen alle helderheidswaarden rechtstreeks voor de gemiddelde oligomere staat van de gedetecteerde eiwitcomplexen worden genormaliseerd. Op dit punt, is het belangrijk om te corrigeren voor de aanwezigheid van niet-belichting fluorescerende FPs die leiden een onderschatting van de oligomere staat tot kan. Dit wordt meestal uitgevoerd door het meten van de helderheid van een homo-dimeric referentie eiwit23,24 met één kleur sFCS of nummer en de helderheid (N & B).
