La production de la plupart des ARN messagers eucaryotes (ARNm) implique l’élimination des introns et la ligature des exons dans un processus nucléaire appelé épissage pré-ARNm1. Deux classes de complexes ARN-protéine (RNP) dirigent le traitement de l’ARN pré-messager en ARNm mature via des complexes splicéosomiques. Une classe, les RNP pré-messagers naissants, est formée co-transcriptionnellement par la liaison de protéines RNP nucléaires hétérogènes et d’autres protéines de liaison à l’ARN, y compris certains membres de la famille SR, produisant des complexes hnRNP2. Les petits RNP nucléaires de deuxième classe, riches en uraciles (SNNP U avec des SNRNA U1, U2, U4, U5 et U6) sont associés à des protéines spécifiques à l’U et au noyau3,4. Les SNNP U interagissent de manière ordonnée avec des régions spécifiques des RNP pré-messagers dans une voie de remodelage dynamique à mesure que les introns sont excisés et que les exons sont ligaturés pour produire des ARNm matures5. De nombreuses protéines nucléaires supplémentaires participent à ces événements de traitement6.
La galectine-1 (Gal1) et la galectine-3 (Gal3) sont deux protéines qui sont des facteurs requis dans la voie d’épissage, comme le montrent les études d’épuisement-reconstitution7,8. L’élimination des deux galectines de l’épissage d’extraits nucléaires compétents (NE) abolit l’assemblage et l’épissage des épissages à un stade précoce. L’ajout de l’une ou l’autre galectine à un NE doublement appauvri restaure les deux activités. Gal1 et Gal3 sont des composants des spliceosomes actifs, comme en témoigne l’immunoprécipitation spécifique du pré-ARNm, des intermédiaires d’épissage et de l’ARNm mature par antisérum spécifique pour Gal1 ou Gal39. Il est important de noter que Gal3 s’associe à des particules endogènes contenant de l’ARN U dans le NE en dehors de la voie d’épissage, comme le montre la précipitation des snRNP par l’antisérum anti-Gal310. Enfin, le silence de Gal3 dans les cellules HeLa modifie les schémas d’épissage de nombreux gènes11.
Dans le NE pré-incubé pour désassembler les spliceosomes préformés12, les snRNP se retrouvent dans de multiples complexes sédimentant dans des gradients de glycérol de 7S à plus de 60S. Bien que le fractionnement du gradient de glycérol soit une technique courante pour l’isolement des complexes et composants splicéosomiques (voir références13,14,15 par exemple), nous avons étendu cette méthode en caractérisant des fractions spécifiques à l’aide d’immunoprécipitations d’anticorps. Un snRNP sédimentant à 10S ne contient que de l’ARNS U1 avec Gal3. L’immunoprécipitation de la fraction 10S avec des antisérums spécifiques pour Gal3 ou U1 snRNP co-précipite à la fois U1 et Gal3 indiquant que certaines des monoparticules SnRNP U1 sont liées à Gal310. Comme U1 snRNP est le premier complexe qui se lie au pré-mRNP dans l’assemblage splicéosomal1,5, cette étape représente un site d’entrée potentiel pour Gal3 dans la voie d’épissage. Sur cette base, nous avons montré que les monoparticules de snRNP 10S Gal3-U1 liées à des perles contenant des anti-Gal3 ont restauré l’activité d’épissage à un NE appauvri en U1 snRNP, établissant ce complexe comme un mécanisme par lequel Gal3 est recruté dans la voie splicéosomale16. Cela contraste avec les tentatives d’isoler les spliceosomes à des stades spécifiques de la réaction d’épissage et de catalogage des facteurs associés17,18. Dans de telles études, la présence de certains facteurs à un moment donné est déterminée, mais pas le mécanisme par lequel ils ont été chargés.
Nous avions précédemment décrit en détail la préparation du NE, le substrat d’épissage, l’assemblage du mélange réactionnel d’épissage et l’analyse des produits dans notre documentation sur le rôle des galectines dans l’épissage pré-ARNm19. Nous décrivons maintenant les procédures expérimentales de fractionnement d’extraits nucléaires pour obtenir une fraction enrichie en complexe snRNP Gal3 – U1 et pour l’immuno-sélection de ce dernier complexe afin de reconstituer l’activité d’épissage dans un extrait nucléaire appauvri en U1.
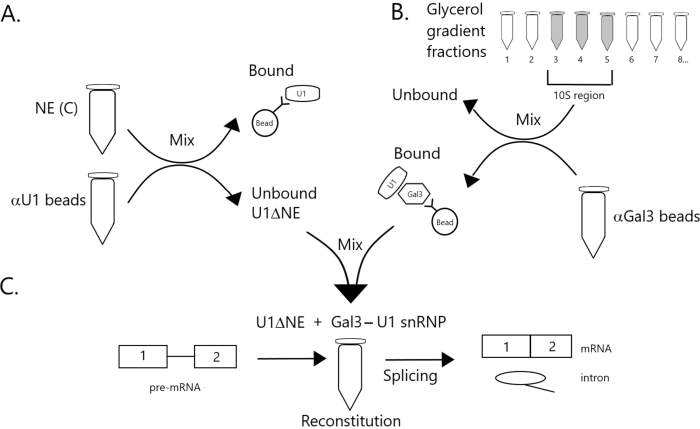
Figure 1 : Schéma illustrant la complémentarité de l’activité d’épissage dans l’extrait nucléaire appauvri en U1 snRNP par un complexe snRNP Gal3-U1 sur des billes. (A) NE dans le tampon C (NE(C)) est incubé avec des billes de protéine A-sépharose couplées de manière covalente avec des billes anti-U1 snRNP (αU1). La fraction non liée est appauvrie en U1 snRNP (U1ΔNE). (B) LE NE dans le tampon D (NE(D)) est fractionné sur un gradient de glycérol de 12 % à 32 % par ultracentrifugation. Les fractions correspondant à la région 10S (fractions 3-5) sont combinées et mélangées avec des billes couplées de manière covalente avec des anticorps anti-Gal3 (billes αGal3). Le matériau lié aux perles contient une monoparticule Gal3-U1 snRNP. (C) Le complexe snRNP Gal3-U1 de la partie (B) est mélangé avec U1ΔNE de la partie (A) dans un essai d’épissage utilisant un substrat pré-ARNm MINX marqué 32P et les intermédiaires et produits de la réaction d’épissage sont analysés par électrophorèse sur gel et autoradiographie. Veuillez cliquer ici pour voir une version agrandie de cette figure.
