Chemical conjugation of αDEC-205 to OVA protein using this protocol will typically allow efficient generation of αDEC-205/OVA for in vivo DC targeting approaches. There are different strategies to verify the technique itself and to test the functionality of the yielded conjugate. Western blot analysis and ELISA are used to verify successful conjugation and at the same time detect potentially left free OVA (Figure 2). In vitro binding studies (Figure 3) and in vivo immunizations (Figure 4) confirm binding of the conjugate to DEC-205 and targeting of DC.
Parallel western blot analysis is used to detect both the conjugated OVA (Figure 2A) as well as conjugated αDEC-205 (Figure 2B). Specifically, the positive signal for OVA at the level of the antibody's molecular weight in the blot confirms association of OVA and the antibody (Figure 2A). Furthermore, staining for OVA in the western blot analysis allows the detection of excess free OVA potentially still present next to the αDEC-205/OVA yielded in step 3.6., which is not the case for the blot shown (Figure 2A). In case large amounts of free OVA are detected, steps 3.4.1. to 3.4.8. of the protocol should be repeated. Complementary to the staining for OVA (Figure 2A), staining for αDEC-205 in western blot analysis verifies successful conjugation through an increase in the molecular weight between "naked" αDEC-205 and the conjugate as shown in Figure 2B.
Next to western blotting, also a specific ELISA allows verification of the successful conjugation of αDEC-205 to OVA. In contrast to the western blot analyses however, this ELISA does not allow the detection of free and un-conjugated αDEC-205 or OVA. Due to the assay setup (Figure 2C), a positive signal is only produced if conjugation was efficient. The positive association between the detected signal (absorption at 450 nm) and the analyzed amount of protein verifies the successful generation of αDEC-205/OVA through chemical conjugation as shown in Figure 2D. At the same time, the positive signal already yielded from 9.38 ng of the αDEC-205/OVA conjugate demonstrates the strong sensitivity of this method (Figure 2D). In case there is no increase in adsorption for increasing amounts of the conjugate, the conjugation was presumably not successful. In this case, also the western blot analyses would yield negative results, i.e., no detection of the conjugate in the blot stained for OVA and no increase in the molecular weight in the blot stained for αDEC-205.
While the western blot and ELISA assays are used to evaluate the conjugation and the removal of free antigen as such, subsequent functional analyses are needed to confirm binding to DEC-205 and targeting of DC. To this end, we have performed in vitro binding studies (Figure 3) and in vivo immunizations (Figure 4). For these experiments, female 6-8 week C57BL/6 and 8-12 week Balb/c mice were obtained from commercial sources or bred at the animal facility of the Helmholtz Centre for Infection Research (HZI) and were housed under specific pathogen-free conditions. Figure 3 demonstrates functional assays for the binding of a conjugate of αDEC-205 and the HCV Core protein (αDEC-205/Core) to CD11c+ cells in vitro. Flow-cytometry clearly showed αDEC-205/Core to efficiently bind bone-marrow derived CD11c+ cells (Figure 3A,B) as well as freshly isolated mouse CD11c+ splenocytes (data not shown). These assays demonstrate the chemical conjugation not to interfere with the binding capacity of αDEC-205. This is further confirmed by immunofluorescence analyses showing binding of αDEC-205/Core to MHC-II+ CD11c+ cells sorted from in vitro generated bone-marrow derived dendritic cells (BMDC) (Figure 3C).
In the past, we have shown the αDEC-205/OVA conjugates produced by the demonstrated protocol to efficiently induce OVA-specific immune responses in vivo in mice, confirming successful generation of the conjugate as well as functional targeting of DC (Figure 4)12,14. Specifically, subcutaneous vaccination with αDEC-205/OVA efficiently induced humoral and cellular OVA-specific immune responses. Importantly, in a recombinant adenovirus challenge model we detected antiviral CD8+ T cells capable of eliminating virus-infected hepatocytes, which has strong implications for vaccines directed at hepatotropic viruses12. Moreover, the highly effective induction of antigen-specific cytotoxic T cells underlines the potential of this approach for the in vivo priming of antitumor immunity. Also, we have successfully used αDEC-205/OVA to test and compare different adjuvants in the context of in vivo DC targeting14. In vaccination with αDEC-205/OVA together with the adjuvant combination Poly(I:C) (polyinosinic-polycytidylic acid) and CpG (synthetic oligodeoxynucleotides containing unmethylated CpG motifs) we observed generally (Figure 4A) and for some time-points significantly higher OVA-specific IgG levels as compared to vaccination with OVA alone (Figure 4B). Furthermore, αDEC-205/OVA efficiently induced OVA-specific CD4+ as well as CD8+ T cell responses (Figure 4C,D) and the αDEC-205/OVA-induced CD8+ T cell response significantly exceeded that induced by OVA alone (Figure 4D).
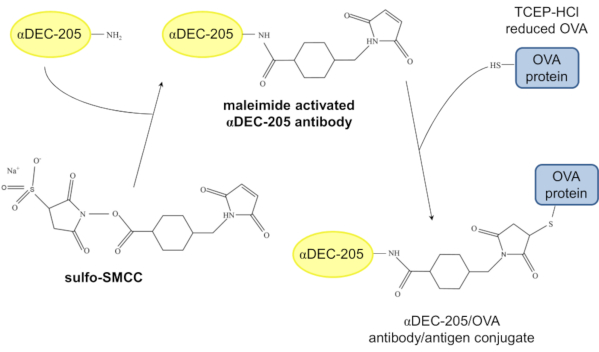
Figure 1: Model of the chemical conjugation of αDEC-205 and OVA. In a first step, the primary amine of αDEC-205 reacts with the NHS ester of the crosslinker sulfo-SMCC resulting in maleimide activated αDEC-205. Following reduction of the disulfide bonds of the OVA protein through incubation with TCEP-HCl, the maleimide activated αDEC-205 reacts with the TCEP-HCl-reduced OVA protein to form the αDEC-205/OVA antibody/antigen conjugate. Abbreviations: N-hydroxysuccinimide ester (NHS ester); sulfosuccinimidyl 4-[N-maleimidomethyl]cyclohexane-1-carboxylate (sulfo-SMCC); Tris(2-carboxysethyl)phosphine hydrochloride (TCEP-HCl). Please click here to view a larger version of this figure.
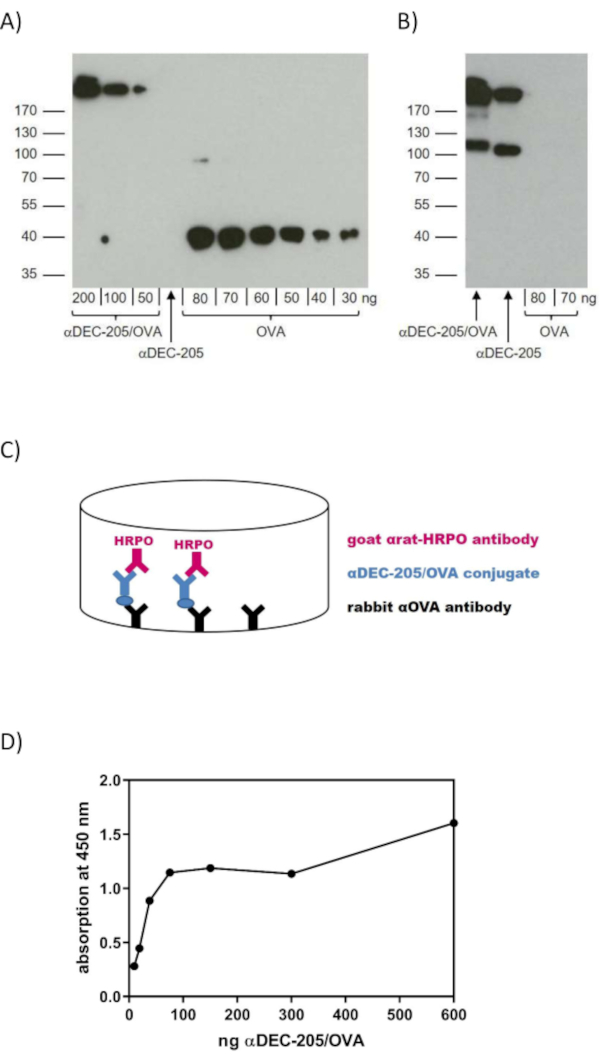
Figure 2: Verification of the chemically conjugated αDEC-205/OVA. To verify effective chemical conjugation of αDEC-205 and the OVA protein, western blot analysis (A,B) and ELISA (C,D) are performed. Samples of αDEC-205/OVA, αDEC-205 and different concentrations of OVA protein were subjected to SDS-PAGE (10%) and subsequent western blot analysis utilizing a rabbit αOVA primary antibody and a goat αrabbit-IgG-HRPO secondary antibody to detect OVA protein (A) or a goat αrat-IgG(H+L)-HRPO antibody to detect αDEC-205 (B). (C) Schematic representation of the ELISA for the verification of the αDEC-205/OVA conjugate. The rabbit αOVA coating antibody binds αDEC-205/OVA via the conjugated OVA. Goat αrat-IgG(H+L)-HRPO recognizes the αDEC-205 fraction of the bound conjugate and a positive signal thus confirms effective conjugation. (D) ELISA was performed as described in (C). Serially diluted amounts (1:2) of αDEC-205/OVA (600 ng to 9.38 ng) were analyzed. Data are shown as the mean of triplicates of a representative assay. Abbreviations: enzyme-linked immunosorbent assay (ELISA); horse radish peroxidase (HRPO); ovalbumin (OVA); sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). Panels A and B have been modified from Volckmar et al.12. http://creativecommons.org/licenses/by/4.0/. Please click here to view a larger version of this figure.
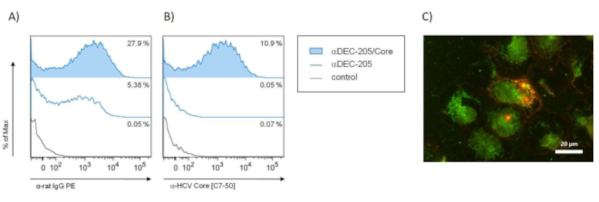
Figure 3: Binding of αDEC-205/Core to bone-marrow derived dendritic cells (BMDC) by flow-cytometry and immunofluorescence microscopy. To analyze the capacity of αDEC-205/Core to bind its target molecule DEC-205 on BMDCs in vitro, fluorescence-activated cell sorting (FACS) analysis (A,B) and immunofluorescence microscopy (C) were performed. In brief, bone marrow cells were isolated from the hind legs of female Balb/c mice (n=3) 8-12 weeks of age and cultured in RPMI medium supplemented with 1% penicillin/streptomycin, 1% glutamine, 0.25 mM mercaptoethanol and 5 ng/mL GM-CSF (granulocyte-macrophage colony stimulating factor). On day 6, the non-adherent BMDCs were carefully harvested and used for binding analysss. (A,B) In vitro generated BMDCs were incubated with 10 µg/mL αDEC-205/Core, αDEC-205 or medium (control) for 1 h at 4 °C followed by staining with APC-labeled αCD11c [clone HL3]. Detection of bound αDEC-205/Core on the surface of BMDCs was performed by additionally staining of the cells with either PE-labeled goat αrat (A) or mouse αHCV Core [clone C7-50] followed by secondary αmouse-IgG1-PE staining (B). Representative histograms show the PE signal and % PE-positive cells of the gated CD11c+ cells. (C) BMDCs generated in vitro from naïve Balb/c mice were sorted for MHC-II+ and CD11c+ cells and were incubated with 10 µg/mL αDEC-205/Core for 1 h at 4 °C. Cell-bound αDEC-205/Core was stained with Alexa 594-coupled αrat-IgG or with mouse αHCV Core [clone C7-50] and Alexa 488-coupled αmouse-IgG for 30 min at 4 °C after washing. The cells were visualized by immunofluorescence microscopy (scale bar = 20 µm). The binding capacity of αDEC-205/Core to BMDCs was confirmed by an overlay of both stainings (double positive = orange). Abbreviations: bone-marrow derived dendritic cells (BMDC); fluorescence-activated cell sorting (FACS); granulocyte-macrophage colony stimulating factor (GM-CSF); hepatitis C virus (HCV). Please click here to view a larger version of this figure.
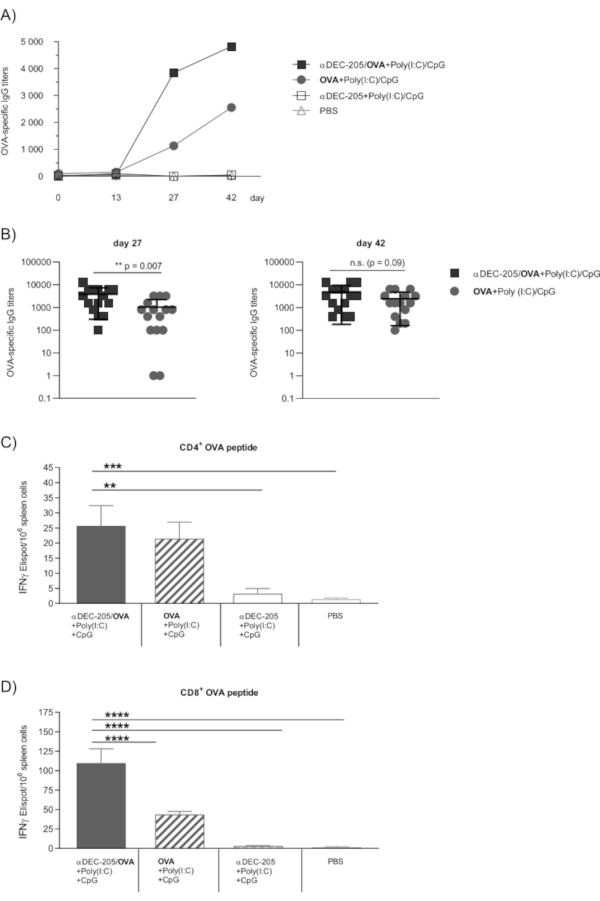
Figure 4: OVA-specific humoral and cellular immune responses following immunization with αDEC-205/OVA. The functionality of αDEC-205/OVA to target DC in vivo was proven through immunization experiments as previously published in Volckmar et al.12. Briefly, female 6-8 week old C57BL/6 mice (n=5) were subcutaneously immunized on days 0, 14 and 28 with 30 µg αDEC-205/OVA conjugate together with the adjuvants 50 µg Poly(I:C)/50 µg CpG, 30 µg αDEC-205 alone or 7 µg OVA protein alone in a total volume of 50 µL PBS per animal. Further controls were treated with PBS alone. (A,B) To monitor the humoral immune response, vaccinated mice were lightly anesthetized through isoflurane inhalation and blood samples were collected from the retro-orbital sinus on day 0, 13 and 27 and by cardiac puncture on day 42. Sera were prepared as described and assayed for the presence of OVA-specific IgG by ELISA12. Endpoint titers were expressed as the reciprocal value of the last serum dilution that yielded an absorbance two times above the values of negative controls. Results are compiled from three independent experiments. (A) Kinetic of OVA-specific total serum IgG titers shown as the group mean. (B) OVA-specific IgG titer on day 27 and day 42 shown for individual mice together with the group mean. Statistics: unpaired two-sided t-test. (C,D) The induction of cellular immune responses was analyzed by enzyme-linked immunosorbent spot (ELISPOT) assays using the murine IFNγ detection kit on day 42 as previously published12. Isolated splenocytes from immunized mice were pooled for the experimental groups and the number of IFNγ spot forming units/106 cells following stimulation with 5 mg/mL CD4+ (C) or CD8+ OVA peptide (D) was analyzed (OVA peptides: CD4323–339 (ISQAVHAAHAEINEAGR) and CD8257–264 (SIINFEKL)). Bars represent the mean ± SEM (n=5, triplicates from pooled samples). Statistics: one-way ANOVA with Dunnett's multiple comparisons test (**p<0.01, ***p<0.001, ****p<0.0001). Abbreviations: cytosine-phosphate-guanine oligonucleotide sequences (CpG), enzyme-linked immunosorbent assay (ELISA), enzyme-linked immunosorbent spot (ELISPOT), ovalbumin (OVA), phosphate-buffered saline (PBS), polyinosinic-polycytidylic acid (Poly(I:C)). This figure has been modified from Volckmar et al.12 http://creativecommons.org/licenses/by/4.0/. Please click here to view a larger version of this figure.
