Les protéines de la superfamille des facteurs de croissance transformants bêta (TGFβ) sont des cytokines pléiotropes avec une variété de membres, y compris TGFβs, protéines morphogénétiques osseuses (BMP) et Activins1,2. La liaison au ligand induit la formation d’oligomères récepteurs conduisant à la phosphorylation et, par conséquent, à l’activation du SMAD régulateur cytosolique (R-SMAD). Selon la sous-famille de ligands, différents R-SMAD sont activés1,2. Alors que les TGFβs et les Activines induisent principalement la phosphorylation de SMAD2/3, les BMP induisent une phosphorylation SMAD1/5/8. Cependant, il y a de plus en plus de preuves que les BMP et les TGFβs/Activins activent également les R-SMAD de l’autre sous-famille respective, dans un processus appelé « signalisation latérale»3,4,5,6,7,8 et qu’il existe des complexes SMAD mixtes composés à la fois de membres SMAD1/5 et SMAD2/333,9 . Deux R-SMAD activés forment par la suite des complexes trimériques avec le médiateur commun SMAD4. Ces complexes de facteurs de transcription sont alors capables de se transloquer dans le noyau et de réguler la transcription des gènes cibles. Les SMAD peuvent interagir avec une variété de co-activateurs et de co-répresseurs transcriptionnels différents, ce qui conduit à la diversification des possibilités de régulation des gènes cibles10. La déréglementation de la signalisation SMAD a de graves implications dans une variété de maladies. Dans cette optique, une signalisation TGFβ/BMP déséquilibrée peut entraîner des pathologies vasculaires graves, telles que l’hypertension artérielle pulmonaire, la télangiectasie hémorragique héréditaire ou l’athérosclérose3,11,12,13,14.
Les cellules endothéliales (CE) forment la couche la plus interne des vaisseaux sanguins et sont donc exposées au stress de cisaillement (SS), une force de friction exercée par le flux visqueux du sang. Fait intéressant, les CE résidant dans les parties du système vasculaire, qui sont exposées à des niveaux élevés de SS laminaires uniformes, sont maintenus dans un état homéostatique et au repos. En revanche, les CE qui présentent des SS faibles et non uniformes, par exemple aux bifurcations ou à la courbure moindre de l’arc aortique, sont prolifératives et activent les voies inflammatoires15. À leur tour, les sites d’EC dysfonctionnels sont enclins à développer l’athérosclérose. Fait intéressant, les CE dans ces zones d’athéroprone affichent des niveaux aberrants élevés de SMAD2/3 et SMAD1/516,17,18 activés. Dans ce contexte, l’amélioration de la signalisation TGFβ/BMP s’est avérée être un événement précoce dans le développement de lésions athérosclérotiques19 et l’interférence avec la signalisation BMP s’est avérée réduire considérablement l’inflammation vasculaire, la formation d’athérome et la calcification associée20.
Le test de ligature de proximité (PLA) est une technique biochimique permettant d’étudier les interactions protéine-protéine in situ21,22. Il repose sur la spécificité des anticorps de différentes espèces qui peuvent se lier aux protéines cibles d’intérêt, permettant une détection très spécifique des interactions protéiques endogènes au niveau d’une seule cellule. Ici, les anticorps primaires doivent se lier à leur épitope cible à une distance inférieure à 40 nm pour permettre la détection23. Par conséquent, le PLA est très bénéfique par rapport aux approches traditionnelles de co-immunoprécipitation, où plusieurs millions de cellules sont nécessaires pour détecter les interactions protéiques endogènes. Dans le PLA, les anticorps secondaires spécifiques à l’espèce, liés de manière covalente à des fragments d’ADN (appelés sondes Plus et Moins), se lient aux anticorps primaires et si les protéines d’intérêt interagissent, les sondes Plus et Moins sont proches. L’ADN est ligaturé à l’étape suivante et l’amplification en cercle roulant de l’ADN circulaire est rendue possible. Lors de l’amplification, des oligonucléotides complémentaires marqués par fluorescence se lient à l’ADN synthétisé, ce qui permet de visualiser ces interactions protéiques par microscopie à fluorescence conventionnelle.
Le protocole décrit ici permet aux scientifiques de comparer quantitativement le nombre de complexes de transcription SMAD actifs dans des conditions athéroprotectrices et athéroprones SS in vitro à l’aide de PLA. SS est généré via un système de pompe pneumatique programmable capable de générer un débit unidirectionnel laminaire de niveaux définis et permettant d’augmenter progressivement les débits. Cette méthode permet de détecter les interactions entre SMAD1/5 ou SMAD2/3 avec SMAD4, ainsi que les complexes mixtes R-SMAD. Il peut facilement être étendu pour analyser les interactions des SMAD avec les co-régulateurs transcriptionnels ou aux complexes de facteurs de transcription autres que les SMAD. La figure 1 montre les principales étapes du protocole présenté ci-dessous.
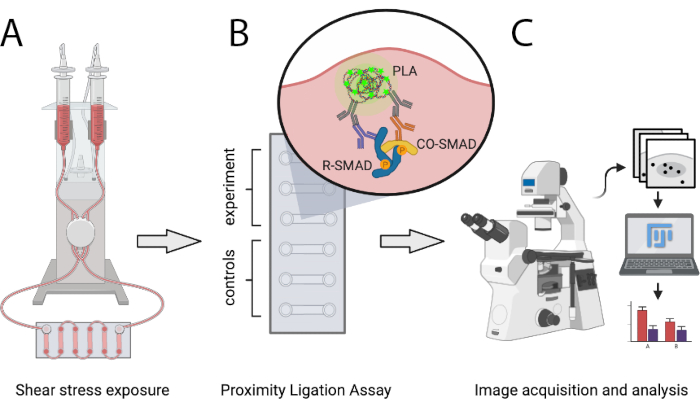
Figure 1 : Représentation schématique du protocole décrit. (A) Les cellules ensemencées dans des lames à 6 canaux sont exposées à une contrainte de cisaillement à l’aide d’un système de pompe pneumatique. (B) Les cellules fixes sont utilisées pour l’expérience du PLA ou pour les conditions de contrôle. (C) Les images des expériences PLA sont acquises avec un microscope à fluorescence et sont analysées à l’aide du logiciel d’analyse ImageJ. Veuillez cliquer ici pour voir une version agrandie de cette figure.
