Salivary glands are relatively easy to dissect from all stage 4 larvae. Male and female larvae can be distinguished at the late L4 larval stage by a red stripe along the dorsal thorax of females but not males (Figure 2). We also observe that antennal morphology is much more elaborate in male than in female L4 larvae (Figure 2), similar to the differences observed in this structure in adult mosquitoes. Along with the considerable overall growth during the L4 stage, the salivary glands also form a lumen during L412. Salivary glands isolated from early L4 stage larvae stained with Hoechst reveal the proximal and distal lobes separated by a narrow constriction (Figure 3B,C). The forming lumen will extend all the way from the salivary duct at the proximal-most end (not shown) through the distal lobe. The forming lumen can be seen in the immunostained distal salivary gland of a mid-to-late L4 larva (Figure 4). The apical domains of the secretory cells surrounding the forming lumen have intense Nile Red staining (Figure 4B,C) suggestive of microvilli-like structures. Also observed close to the apical surface is Rab11 staining (Figure 4C,D, arrows). Rab11 localizes to apical recycling endosomes. The Rab11 staining that also accumulates along the basal surface of the gland is an artefact due to the stickiness of the basal membrane. Similar background staining is common with immunostaining of both larval and adult salivary glands and has been mistaken for bona fide signal.
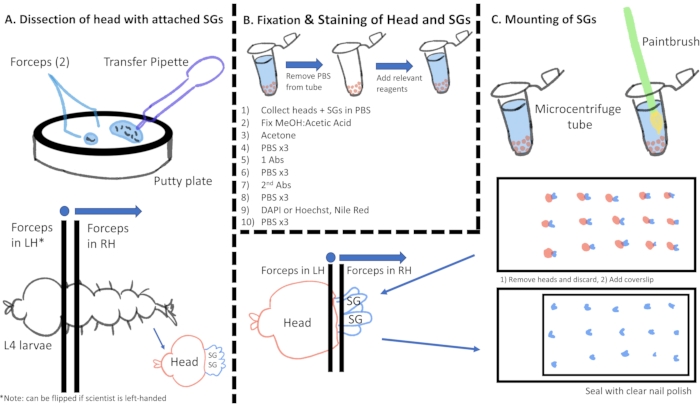
Figure 1: Cartoon visualization of the immunostaining process from gland dissection through slide preparation. Abbreviations: SGs = salivary glands; PBS = phosphate-buffered saline; MeOH = methanol; DAPI = 4′,6-diamidino-2-phenylindole; Abs = antibodies; LH = left hand; RH = right hand. Please click here to view a larger version of this figure.

Figure 2: Male and female L4 stage Anopheles gambaie larvae. (A, B) Early L4 larvae. (C, D) Late L4 Larvae. There is considerable growth during the L4 stage. Females (A, C) have been described as having a distinguishing red stripe down the dorsal thorax (C; black arrow) that is not present in males (B, D), but also their antennae are much less elaborate (frilly) than those of male larvae from both early and late L4 (white arrows in enlarged images). Scale bars = 1 mm. Please click here to view a larger version of this figure.
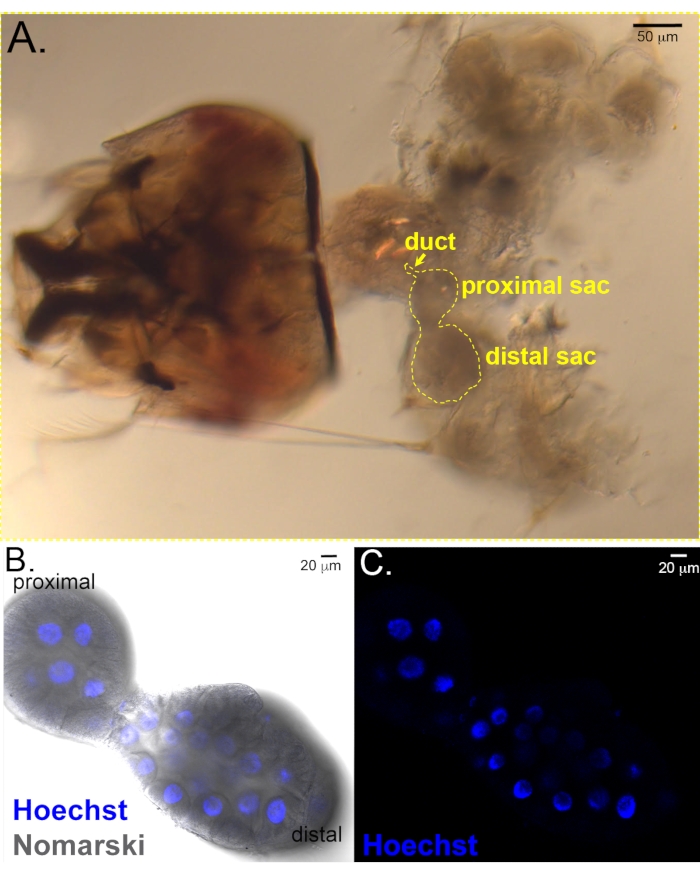
Figure 3: Early L4 salivary glands. (A) Early L4 female larval head with salivary glands and other internal organs still attached. Salivary gland is outlined. Scale bar = 50 µm. (B, C) Isolated larval glands stained with Hoechst. (B) Merge of fluorescent and Nomarski images showing the shape of the proximal and distal lobes and the blue Hoechst staining in nuclei. (C) Hoechst fluorescent image highlights nuclei only. Scale bar in bottom panels = 20 µm. Please click here to view a larger version of this figure.
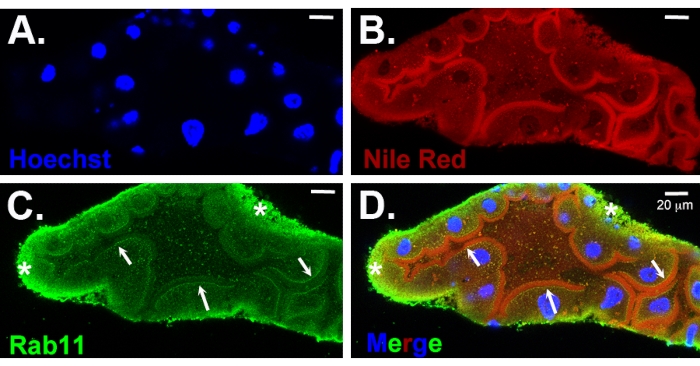
Figure 4: Immunostained distal L4 salivary gland. (A) Gland has been stained with Hoechst (nuclear DNA, blue) and (B) Nile Red (membrane marker, red); immunostained for Rab11 (apical recycling vesicles, green) (C). (D) The merged image. Note that a lumen is present at the stage shown (B–D). The arrows in C and D point to the expected Rab11 staining of vesicles near the apical surface that overlaps the Nile Red-positive vesicular staining in the merge (white arrows). Non-specific background staining around the basal surface (asterisks) is also observed, a common type of background observed in larval and adult salivary glands. Scale bars = 20 µm. Please click here to view a larger version of this figure.
