成骨细胞形成的骨骼伴随着破骨细胞对骨骼的协调破坏或再吸收。成骨细胞骨形成和破骨细胞再吸收之间的平衡是描述骨转换或重塑的耦合过程,这对于骨骼稳态至关重要。成骨细胞功能障碍导致骨骼形成受损并导致各种疾病,包括骨质疏松症1,2,3。骨髓基质干细胞(BMSCs)到成骨细胞前体和成熟成骨细胞的 离体/体外 分化导致矿化骨基质随着时间的推移在培养血管中形成和沉积4,5,6。成骨细胞形成的这种骨骼需要大量的细胞能量。具体而言,胶原蛋白的合成和分泌已被证明严重依赖于细胞ATP:ADP比率,并且可能,矿化囊泡运输和分泌需要额外的ATP7,8,9,10,11。许多研究人员已经证明,成骨细胞生成和成骨细胞功能的过程需要足够的能量供应来满足骨形成的代谢需求12,13,14,15,16。因此,该方法的目标是使用实时细胞代谢通量分析仪表征原代,小鼠基质细胞在整个成骨细胞分化过程中的生物能量状态。这些技术有助于更好地了解骨骼稳态,这可能最终导致能够改善骨骼疾病的新型治疗方案的发展。
实时细胞代谢通量分析仪可用于测量活成骨细胞的耗氧速率(OCR)和细胞外酸化速率(ECAR),分别对应于线粒体和糖酵解ATP的产生。这种方法的基础是,在糖酵解过程中,每个乳酸释放一个H + 离子,将葡萄糖转化为乳酸盐,这会改变ECAR值中反映的培养基pH值。相反,在TCA(三羧酸)循环期间, 通过 线粒体的氧化磷酸化通过利用或消耗氧气产生CO2 ,因此监测OCR反映了这种代谢过程。该分析仪同时实时测量细胞外微环境中的OCR和ECAR,这在研究细胞生物能量学时具有巨大的潜力6,17。此外,进行这些测定相对简单,并且易于根据实验目标进行定制。已经采用类似的技术来进一步了解免疫系统的T细胞代谢调节18,19,癌症的发生和进展20,以及导致代谢综合征的多种其他细胞类型21,22。
与替代技术相比,实时代谢通量分析仪的优势包括:(1)能够实时测量活细胞的细胞生物能量,(2)能够对相对较少的细胞进行测定(需要低至5,000个细胞),(3)注射端口在高通量96孔系统中并行操作多个处理,(4)使用无放射性标记的自动细胞成像仪进行归一化18,23,24.以下方法旨在提供使用分析仪在整个成骨细胞分化过程中监测小鼠BMSC中细胞生物能量学的广义但详细的描述。它将包括常规进行的测定;然而,与许多技术和方法一样,强烈建议各个实验室确定其实验的具体细节。
可选择测定和不同类型的测定:提供多种测定试剂盒和试剂,用于研究细胞的生物能量学,同时确保实验结果的可靠性和一致性。此外,桌面软件还提供可轻松定制的分析模板。该测定可以根据用户测量不同代谢参数的需要来定义。这些测定可以根据实验目标和/或科学问题以各种方式进行修改。例如,使用四个进样端口,可以将多种化合物注射到测定培养基中,以分析特定于每个代谢途径的细胞反应。
细胞能量表型测试: 该测定法测量活细胞的代谢表型和代谢潜力。该测定也被推荐作为获得通路特异性代谢的广义概念的第一步。注入寡霉素A-ATP合酶抑制剂和羰基氰化物4-(三氟甲氧基)苯腙(FCCP)-线粒体解偶联剂的混合物以了解细胞能量电位。注射寡霉素A抑制ATP的合成,导致糖酵解速率(ECAR)的增加,使细胞能够满足其能量需求;另一方面,由于线粒体膜的去极化,FCCP的注射导致更高的OCR。从本质上讲,该测定描绘了基础代谢呼吸,并在双重注射,推或应激之后,代谢反应。基于这些参数,软件然后通过将细胞分类为有氧,静止,糖酵解或能量状态来绘制细胞的OCR和ECAR,时间跨度25,26。
ATP实时产率测定: 这测量细胞ATP同时从糖酵解和线粒体呼吸产生的。该测定定量测量来自两个能量途径的代谢变化,并提供有关线粒体和糖酵解ATP产生速率随时间变化的数据。该测定获得基础OCR和ECAR数据,然后通过注射寡霉素A计算线粒体ATP产生速率,通过注射鱼藤酮+抗霉素A混合物计算糖酵解ATP产生速率(完全抑制线粒体功能),导致线粒体酸化17,27。
细胞线粒体负荷试验(或细胞丝粒体负荷试验): 这通过ATP连接的呼吸测量线粒体功能,量化细胞生物能量,识别线粒体功能障碍,并测量细胞对压力的反应。一次测定即可获得各种参数,包括基础和备用呼吸能力、ATP 相关呼吸、最大呼吸和非线粒体耗氧量。该测定涉及序贯注射寡霉素A,FCCP(线粒体解偶联剂),鱼藤酮/抗霉素A抑制剂的混合物,以有效分析这些抑制剂对线粒体功能的影响28。
柔韧性水母燃料柔性测试: 这通过三种主要线粒体燃料的存在和不存在抑制剂的氧化来测量线粒体呼吸速率。葡萄糖,谷氨酰胺和脂肪酸的顺序抑制有助于测量细胞的依赖性,容量和柔韧性以及细胞在各种细胞途径中的依赖性,以满足能量需求。当线粒体不能通过氧化其他燃料来满足阻塞的靶向途径的需求时,细胞进入依赖状态。细胞的容量是通过抑制其他两种替代途径,然后抑制感兴趣的途径来计算的。细胞的灵活性有助于了解线粒体补偿和满足抑制途径的燃料需求的能力。它是通过从单元的容量中减去单元的依赖性来计算的。独立使用三种不同的抑制剂或作为两种抑制剂的混合物来有效计算测定参数。2-氰基-3-(1-苯基-1H-吲哚-3-基)-2-丙烯酸(UK5099)通过阻断糖酵解中的丙酮酸盐载体来抑制葡萄糖的氧化。双-2-(5-苯基乙酰氨基-1,3,4-噻二唑-2-基)(BPTES)乙基硫醚抑制谷氨酰胺氧化途径,依托莫西抑制长链脂肪酸的氧化29。
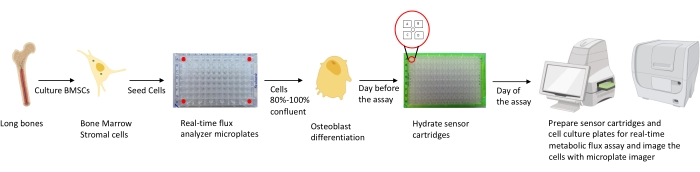
图1:用于培养和制备成骨细胞进行分析的方法示意图。 从长骨中分离出小鼠BMSC,培养并接种在96孔板中,密度为25,000个细胞/ 孔。当这些细胞达到80%-100%汇合以开始分化时,就开始在成骨细胞特异性培养基中培养这些细胞。测定在不同的分化阶段进行。在测定前一天将筒板水合。在测定当天,根据测定要求将不同的抑制剂注射到传感器盒的端口中,并将校准缓冲液添加到96孔校准板中。校准后,进行实时细胞代谢通量测定,然后使用微孔板成像仪对细胞培养微孔板进行成像,以使用细胞计数对实时细胞代谢通量分析仪数据进行归一化。 请点击此处查看此图的大图。
