De vorming van bot door de osteoblast gaat gepaard met gecoördineerde vernietiging of resorptie van botten door osteoclasten. De balans tussen osteoblastische botvorming en osteoclastenresorptie is een gekoppeld proces dat botomzetting of remodellering beschrijft, wat essentieel is voor skelethomeostase. Osteoblastdisfunctie leidt tot verminderde botvorming en resulteert in verschillende ziekten, waaronder osteoporose 1,2,3. Ex vivo/in vitro differentiatie van beenmergstromale stamcellen (BMSCs) naar osteoblastvoorlopers en volwassen osteoblasten resulteert in de vorming en afzetting van de gemineraliseerde botmatrix in het kweekvat in de loop van de tijd 4,5,6. Deze botvorming door de osteoblast vereist een aanzienlijke hoeveelheid cellulaire energie. In het bijzonder is aangetoond dat collageensynthese en secretie sterk afhankelijk zijn van cellulaire ATP: ADP-verhoudingen, en vermoedelijk vereisen gemineraliseerde blaasjeshandel en secretie extra ATP 7,8,9,10,11. Veel onderzoekers hebben aangetoond dat het proces van osteoblastogenese en osteoblastfunctie een voldoende toevoer van energie vereist om te voldoen aan de metabole vraag naar botvorming 12,13,14,15,16. Daarom is het doel van deze methode om de bio-energetische status van primaire, muriene stromale cellen te karakteriseren gedurende osteoblastdifferentiatie met behulp van de real-time cel metabole flux analyzer. Deze technieken helpen bij het ontwikkelen van een beter begrip van skelethomeostase, wat uiteindelijk kan leiden tot de ontwikkeling van nieuwe therapeutische opties die skeletaandoeningen kunnen verbeteren.
De real-time cel metabole flux analyzer kan worden gebruikt om de zuurstofverbruikssnelheid (OCR) en extracellulaire verzuringssnelheid (ECAR) van levende osteoblasten te meten, wat overeenkomt met respectievelijk mitochondriale en glycolytische ATP-productie. Fundamenteel voor deze methodologie is het feit dat één H + –ion per lactaat vrijkomt tijdens glycolyse bij de omzetting van glucose in lactaat, wat de media-pH verandert die wordt weerspiegeld in de ECAR-waarden. Omgekeerd, tijdens de TCA (tricarbonzuur) cyclus, oxidatieve fosforylering via de mitochondriën produceert CO2 door zuurstof te gebruiken of te consumeren, en daarom is het monitoren van OCR een weerspiegeling van dit metabolische proces. De analyzer meet zowel OCR als ECAR in de extracellulaire micro-omgeving tegelijkertijd en in realtime, wat een enorm potentieel mogelijk maakt bij het bestuderen van cellulaire bio-energetica 6,17. Bovendien is het uitvoeren van deze testen relatief eenvoudig en gemakkelijk aan te passen, afhankelijk van het experimentele doel. Vergelijkbare technieken zijn gebruikt om de metabool regulatie van het immuunsysteem18,19, kankerinitiatie en progressie20 verder te begrijpen, samen met meerdere andere celtypen die bijdragen aan metabool syndroom21,22.
De voordelen van real-time metabole flux analyzer ten opzichte van alternatieve technieken omvatten (1) de mogelijkheid om cellulaire bio-energetica van levende cellen in real-time te meten, (2) het vermogen om assay uit te voeren met een relatief klein aantal cellen (vereist slechts 5.000 cellen), (3) injectiepoorten om parallel meerdere behandelingen te manipuleren in een high-throughput 96-well systeem, (4) gebruik van radioactieve labelvrije geautomatiseerde celbeeldcamera voor normalisatie18, 23,24. De volgende methoden zijn bedoeld om een algemene maar gedetailleerde beschrijving te geven van het monitoren van cellulaire bio-energetica in muriene BMSCs gedurende de differentiatie van osteoblasten met behulp van de analyzer. Het omvat routinematig uitgevoerde assays; echter, zoals met veel technieken en methoden, wordt het sterk aangemoedigd dat individuele laboratoria specifieke details voor hun experimenten bepalen.
Selectie van assays en verschillende soorten assays beschikbaar: Een breed scala aan assay kits en reagentia zijn beschikbaar om de bio-energetica van cellen te bestuderen en tegelijkertijd de betrouwbaarheid en consistentie van de experimentele resultaten te waarborgen. Bovendien biedt de desktopsoftware ook testsjablonen die eenvoudig kunnen worden aangepast. De test kan worden gedefinieerd op basis van de behoeften van de gebruiker om verschillende metabole parameters te meten. Deze assays kunnen op verschillende manieren worden aangepast op basis van het experimentele doel en/of de wetenschappelijke vraag. Met vier injectiepoorten kunnen bijvoorbeeld meerdere verbindingen in de testmedia worden geïnjecteerd om de cellulaire respons te analyseren die specifiek is voor elke metabole route.
Celenergie fenotype test: Deze test meet het metabole fenotype en het metabolisch potentieel van de levende cellen. Deze test wordt ook aanbevolen als de eerste stap om een gegeneraliseerd idee te krijgen van pathway-specifiek metabolisme. Een mengsel van oligomycine A-een remmer van ATP-synthase en carbonylcyanide 4-(trifluormethoxy) fenylhydrazon (FCCP)-een mitochondriaal ontkoppelingsmiddel wordt geïnjecteerd om het celenergiepotentieel te begrijpen. De injectie van oligomycine A remt de synthese van ATP, wat resulteert in een toename van de snelheid van glycolyse (ECAR) om de cellen in staat te stellen aan hun energiebehoeften te voldoen; aan de andere kant resulteert de injectie van FCCP in een hogere OCR als gevolg van depolarisatie van het mitochondriale membraan. In wezen toont deze test basale metabole ademhaling en na de dubbele injecties, duwen of spanningen de metabole respons. Op basis van deze parameters plot de software vervolgens OCR en ECAR van de cellen door de cellen te classificeren als aerobe, rustige, glycolytische of energetische toestand in de loop van de tijd25,26.
ATP real-time productiesnelheidstest: Dit meet de cellulaire ATP-productie tegelijkertijd uit glycolyse en mitochondriale ademhaling. Deze test meet kwantitatief de metabole verschuivingen van de twee energieroutes en levert gegevens over de mitochondriale en glycolytische ATP-productiesnelheden in de loop van de tijd. De test verkrijgt basale OCR- en ECAR-gegevens, gevolgd door het berekenen van mitochondriale ATP-productiesnelheid door injectie van oligomycine A en glycolytische ATP-productiesnelheid door injectie van rotenon + antimycine A-mengsel (totale remming van mitochondriale functie), resulterend in mitochondriale verzuring17,27.
Cel mitochondriën stresstest (of cel mito stress test): Dit meet de mitochondriale functie door atp-gebonden ademhaling, kwantificeert cellulaire bio-energetica, identificeert mitochondriale disfunctie en meet de reactie van cellen op stress. Verschillende parameters, waaronder basale en reserve ademhalingscapaciteit, ATP-gebonden ademhaling, maximale ademhaling en niet-mitochondriaal zuurstofverbruik, kunnen in één test worden verkregen. Deze test omvat sequentiële injecties van oligomycine A, FCCP (mitochondriaal ontkoppelingsmiddel), een mengsel van rotenon / antimycine A-remmers om het effect hiervan op de mitochondriale functie efficiënt te analyseren28.
Flexibiliteit mito fuel flex test: Dit meet de mitochondriale ademhalingssnelheid door de oxidatie van de drie primaire mitochondriale brandstoffen door de aan- en afwezigheid van hun remmers. De sequentiële remming van glucose, glutamine en vetzuren helpt bij het meten van de afhankelijkheid, capaciteit en flexibiliteit van cellen en de afhankelijkheid van de cellen in verschillende cellulaire routes om aan de energievraag te voldoen. Wanneer de mitochondriën niet kunnen voldoen aan de eisen van de geblokkeerde route van belang door andere brandstoffen te oxideren, komen de cellen in een afhankelijkheidstoestand. De capaciteit van de cellen wordt berekend door remming van de andere twee alternatieve routes gevolgd door de remming van de route van belang. De flexibiliteit van cellen helpt bij het begrijpen van het vermogen van mitochondriën om te compenseren en te voldoen aan de brandstofbehoeften van de geremde route. Het wordt berekend door de afhankelijkheid van cellen af te trekken van de capaciteit van cellen. Drie verschillende remmers worden onafhankelijk van elkaar of als een mengsel van twee gebruikt om de testparameters effectief te berekenen. 2-cyano-3-(1-fenyl-1H-indol-3-yl)-2-propeenzuur (UK5099) remt de oxidatie van glucose door de pyruvaatdrager in glycolyse te blokkeren. Bis-2-(5-fenylacetamido-1,3,4-thiadiazol-2-yl) (BPTES) ethylsulfide remt de glutamine-oxidatieroute en etomoxir remt de oxidatie van langeketenvetzuren29.
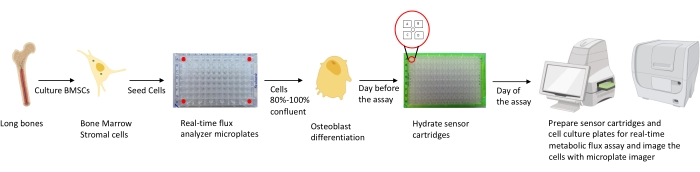
Figuur 1: Schematische weergave van de methodologie voor het kweken en voorbereiden van osteoblasten voor analyse. Murine BMSC’s worden geïsoleerd uit lange botten, gekweekt en gezaaid in 96-well platen met 25.000 cellen / putdichtheid. Het kweken van deze cellen in osteoblastspecifieke media wordt gestart wanneer ze 80% -100% confluentie bereiken om hun differentiatie te starten. De assays worden uitgevoerd in verschillende stadia van differentiatie. De patroonplaten worden een dag voorafgaand aan de test gehydrateerd. Op de dag van de test worden verschillende remmers geïnjecteerd in de poorten van de sensorcartridges op basis van de testvereisten en wordt een kalibratiebuffer toegevoegd aan de 96-well kalibratieplaat. Na kalibratie wordt de real-time cel metabole flux assay uitgevoerd, gevolgd door het in beeld brengen van de celkweekmicroplaat met behulp van de microplate imager om real-time cel metabole flux analyzer gegevens met celgetal te normaliseren. Klik hier om een grotere versie van deze figuur te bekijken.
