Dannelsen av bein av osteoblasten er ledsaget av koordinert ødeleggelse eller resorpsjon av bein av osteoklaster. Balansen mellom osteoblastisk bendannelse og osteoklast resorpsjon er en koblet prosess som beskriver beinomsetning eller ombygging, noe som er viktig for skjelett homeostase. Osteoblast dysfunksjon fører til nedsatt bendannelse og resulterer i ulike sykdommer, inkludert osteoporose 1,2,3. Ex vivo/in vitro differensiering av benmargsstromale stamceller (BMSCer) til osteoblastforløpere og modne osteoblaster resulterer i dannelse og avsetning av den mineraliserte beinmatrisen i kulturfartøyet over tid 4,5,6. Denne beindannelsen ved osteoblasten krever en betydelig mengde cellulær energi. Spesielt har kollagensyntese og sekresjon vist seg å stole sterkt på cellulær ATP: ADP-forhold, og antagelig krever mineralisert vesicle trafficking og sekresjon ytterligere ATP 7,8,9,10,11. Mange forskere har vist at prosessen med osteoblastogenese og osteoblastfunksjon krever tilstrekkelig tilførsel av energi for å møte den metabolske etterspørselen etter beindannelse 12,13,14,15,16. Derfor er målet med denne metoden å karakterisere den bioenergetiske statusen til primære, murine stromale celler gjennom osteoblastdifferensiering ved hjelp av sanntidscellemetabolisk fluksanalysator. Disse teknikkene bidrar til å utvikle en bedre forståelse av skjelett homeostase, noe som til slutt kan føre til utvikling av nye terapeutiske alternativer som er i stand til å forbedre skjelettsykdommer.
Sanntidscellemetabolismens fluksanalysator kan brukes til å måle oksygenforbruket (OCR) og ekstracellulær forsuringsrate (ECAR) av levende osteoblaster, som tilsvarer henholdsvis mitokondrie- og glykolytisk ATP-produksjon. Grunnleggende for denne metoden er det faktum at en H + ion per laktat frigjøres under glykolyse i konvertering av glukose til laktat, noe som endrer media pH reflektert i ECAR-verdiene. Motsatt, under TCA (trikarboksylsyre) syklus, oksidert fosforylering via mitokondriene produserer CO2 ved å bruke eller konsumere oksygen, og derfor overvåking OCR reflekterer denne metabolske prosessen. Analysatoren måler både OCR og ECAR i det ekstracellulære mikromiljøet samtidig og i sanntid, noe som gir et enormt potensial når man studerer cellulær bioenergi 6,17. I tillegg er det relativt enkelt å utføre disse analysene, avhengig av det eksperimentelle målet. Lignende teknikker har blitt brukt for å forstå T-celle metabolsk regulering av immunsystemet18,19, kreftinitiering og progresjon20, sammen med flere andre celletyper som bidrar til metabolske syndromer 21,22.
Fordelene med sanntids metabolsk fluksanalysator over alternative teknikker inkluderer (1) evnen til å måle cellulær bioenergigetikk av levende celler i sanntid, (2) evne til å utføre analyser med et relativt lite antall celler (krever så lite som 5000 celler), (3) injeksjonsporter for å parallelt manipulere flere behandlinger i et 96-brønnssystem med høy gjennomstrømning, (4) bruk av radioaktiv etikettfri automatisert celleavbildning for normalisering18, 23,24. Følgende metoder tar sikte på å gi en generalisert, men detaljert beskrivelse av overvåking av cellulær bioenergi i murine BMSCer gjennom osteoblastdifferensiering ved hjelp av analysatoren. Det vil inkludere rutinemessig utførte analyser; Men som med mange teknikker og metoder, er det sterkt oppmuntret at individuelle laboratorier bestemmer spesifikke detaljer for sine eksperimenter.
Utvalg av analyser og ulike typer analyser tilgjengelig: Et bredt utvalg av analysesett og reagenser er tilgjengelige for å studere bioenergien til celler samtidig som påliteligheten og konsistensen av de eksperimentelle resultatene sikres. I tillegg tilbyr skrivebordsprogramvaren også analysemaler som enkelt kan tilpasses. Analysen kan defineres basert på brukerens behov for å måle ulike metabolske parametere. Disse analysene kan endres på ulike måter basert på det eksperimentelle målet og/eller det vitenskapelige spørsmålet. For eksempel, med fire injeksjonsporter, kan flere forbindelser injiseres i analysemediet for å analysere den cellulære responsen som er spesifikk for hver metabolske bane.
Celleenergi fenotype test: Denne analysen måler levende cellers metabolske fenotype og metabolske potensial. Denne analysen anbefales også som det første trinnet for å få en generalisert ide om banespesifikk metabolisme. En blanding av oligomycin A-an-hemmer av ATP-syntase og Carbonylcyanid 4-(trifluorometoksy) fenylhydrazon (FCCP)-et mitokondrie-frakoblingsmiddel injiseres for å forstå celleenergipotensialet. Injeksjonen av oligomycin A hemmer syntesen av ATP, noe som resulterer i en økning i glykolysehastigheten (ECAR) for å gjøre det mulig for cellene å møte sine energibehov; På den annen side resulterer injeksjonen av FCCP i høyere OCR på grunn av depolarisering av mitokondriemembranen. I hovedsak skildrer denne analysen basal metabolsk åndedrett, og etter de doble injeksjonene, pressene eller stressene, den metabolske responsen. Basert på disse parametrene plotter programvaren deretter OCR og ECAR av cellene ved å klassifisere cellene som aerob, passiv, glykolytisk eller energisk tilstand over tid25,26.
ATP-analyse i sanntidsproduksjonsrate: Dette måler den cellulære ATP-produksjonen samtidig fra glykolyse og mitokondrie respirasjon. Denne analysen måler kvantitativt de metabolske skiftene fra de to energiveiene og gir data om mitokondrie- og glykolytiske ATP-produksjonsrater over tid. Analysen innhenter basale OCR- og ECAR-data etterfulgt av beregning av mitokondrie-ATP-produksjonshastighet gjennom injeksjon av oligomycin A og glykolytisk ATP-produksjonshastighet gjennom injeksjon av rotenon + antimycin En blanding (total hemming av mitokondriefunksjon), noe som resulterer i mitokondrieforsuring17,27.
Celle mitokondrier stress test (eller celle mitokon stress test): Dette måler mitokondriefunksjonen gjennom ATP-koblet respirasjon, kvantifiserer cellulær bioenergi, identifiserer mitokondrie dysfunksjon og måler cellenes respons på stress. Ulike parametere, inkludert basal og ekstra respiratorisk kapasitet, ATP-koblet åndedrett, maksimal respirasjon og ikke-mitokondrie oksygenforbruk, kan oppnås i en analyse. Denne analysen innebærer sekvensielle injeksjoner av oligomycin A, FCCP (mitokondrie-frakoblingsmiddel), en blanding av rotenon/antimycin A-hemmere for effektivt å analysere effekten av disse på mitokondriefunksjonen28.
Fleksibilitet mito drivstoff flex test: Dette måler mitokondrie respirasjonshastigheten ved oksidasjon av de tre primære mitokondriebrenselene ved tilstedeværelse og fravær av deres inhibitorer. Den sekvensielle hemmingen av glukose, glutamin og fettsyrer hjelper til med å måle avhengigheten, kapasiteten og fleksibiliteten til celler og avhengigheten av cellene i ulike cellulære veier for å møte energibehovet. Når mitokondriene ikke kan oppfylle kravene til den blokkerte veien av interesse ved å oksidere andre drivstoff, går cellene inn i en avhengighetstilstand. Kapasiteten til cellene beregnes ved hemming av de to andre alternative banene etterfulgt av hemming av interesseveien. Fleksibiliteten til celler bidrar til å forstå mitokondrienes evne til å kompensere og møte drivstoffbehovene til den hemmede banen. Den beregnes ved å trekke avhengigheten av celler fra cellekapasiteten. Tre forskjellige hemmere brukes uavhengig eller som en blanding av to for effektivt å beregne analyseparametrene. 2-cyano-3-(1-fenyl-1H-indol-3-yl)-2-propenoic acid (UK5099) hemmer oksidasjon av glukose ved å blokkere pyruvatbæreren i glykolyse. Bis-2-(5-fenylacetamido-1,3,4-tiadiazol-2-yl) (BPTES) etylsulfid hemmer glutaminoksidasjonsveien, og etomoksir hemmer oksidasjon av langkjedede fettsyrer29.
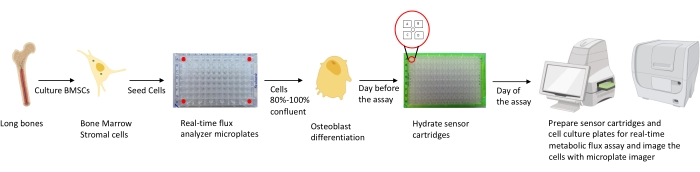
Figur 1: Skjematisk fremstilling av metodikken for dyrking og tilberedning av osteoblaster for analyse. Murine BMSC-er er isolert fra lange bein, dyrket og frøet i 96-brønnsplater ved 25 000 celler / brønntetthet. Kultivering av disse cellene i osteoblastspesifikke medier startes når de når 80% -100% samløp for å starte differensiering. Analysene utføres på forskjellige stadier av differensiering. Patronplatene er hydrert en dag før analysen. På analysedagen injiseres forskjellige inhibitorer i portene til sensorpatronene basert på analysekravene, og en kalibreringsbuffer legges til 96-brønns kalibreringsplaten. Etter kalibrering utføres sanntidscellemetabolismens fluksanalyse, etterfulgt av avbildning av cellekulturmikroplaten ved hjelp av mikroplateavbildningen for å normalisere sanntidscellemetaboliske fluksanalysatordata med celleantall. Klikk her for å se en større versjon av denne figuren.
