De caspasen zijn een familie van cysteïne aspartaat proteasen die kunnen worden gegroepeerd in initiator caspasen en beul caspasen. Beul caspasen bestaan uit caspase-3, -6 en -7. Ze komen van nature voor in cellen als dimeren en worden door de initiator caspasen gesplitst om apoptose1 uit te voeren. Initiator caspasen omvatten human caspase-1, -2, -4, -5, -8, -9, -10 en -12. Ze worden gevonden als inactieve zymogenen (pro-caspasen) die worden geactiveerd door nabijheidsgeïnduceerde dimerisatie en gestabiliseerd door auto-proteolytische splitsing 2,3. De inflammatoire caspasen zijn een subset van de initiator caspasen2 en omvatten caspase-1, -4, -5 en -12 bij mensen, en caspase-1, -11 en -12 bij muis 4,5. In plaats van een apoptotische rol, spelen ze een centrale rol bij ontstekingen. Ze bemiddelen proteolytische verwerking en secretie van pro-interleukine (IL)-1β en pro-IL-18 6,7, de eerste cytokines die vrijkomen als reactie op pathogene indringers 8,9. Caspase-1 wordt geactiveerd bij werving op zijn activeringsplatform; een eiwitcomplex met een groot molecuulgewicht dat het inflammasoom wordt genoemd (figuur 1A)10. Dimerisatie van caspase-4, -5 en -11 vindt onafhankelijk van deze platforms plaats via een niet-canonieke inflammasoomroute11,12.
Canonieke inflammasomen zijn cytosolische multimere eiwitcomplexen die bestaan uit een inflammasoomsensoreiwit, het adaptoreiwit ASC (apoptose-geassocieerd speck-achtig eiwit dat een CARD bevat) en het effectoreiwit caspase-110. De best bestudeerde canonieke inflammasomen zijn de NOD-achtige receptorfamilie met een pyrinedomein (NLRP), NLRP1 en NLRP3, de NLR-familie met een CARD (NLRC), NLRC4 en de afwezige in melanoom 2 (AIM2). Ze bevatten elk een pyrinedomein, een CARD of beide domeinen. Het CARD-domein bemiddelt de interactie tussen CARD-bevattende caspasen en hun upstream activators. Daarom is het steigermolecuul ASC, dat bestaat uit een N-terminal pyrinedomein (PYD) en een C-terminal CARD-motief13,14, vereist voor de rekrutering van caspase-1 voor deNLRP1 10, NLRP315 en AIM216 inflammasomen.
Elk inflammasoom is vernoemd naar zijn unieke sensoreiwit dat verschillende pro-inflammatoire stimuli herkent (figuur 1B). Activatoren van deze route worden canonieke stimuli genoemd. Inflammasomen dienen als sensoren voor microbiële componenten en weefselstress en assembleren om een robuuste ontstekingsreactie te veroorzaken door activering van de inflammatoire caspasen17. Inflammasoomassemblage initieert caspase-1-activering om rijping en secretie van de belangrijkste substraten pro-IL-1β en pro-IL-18 te bemiddelen. Dit proces verloopt via een mechanisme in twee stappen. Ten eerste upreguleert een priming stimulus de expressie van bepaalde inflammasoomeiwitten en pro-IL-1β door activering van de NF-κB-route. Ten tweede induceert een intracellulaire (canonieke) stimulus inflammasoomassemblage en rekrutering van procaspase-1 6,7.
Caspase-4 en caspase-5 zijn de menselijke orthologen van murine caspase-1111. Ze worden op een inflammasoomonafhankelijke manier geactiveerd door intracellulair lipopolysaccharide (LPS), een molecuul dat wordt aangetroffen in het buitenmembraan van Gram-negatieve bacteriën 18,19,20, en door extracellulair heem, een product van hemolyse van rode bloedcellen 21. Er is voorgesteld dat LPS zich rechtstreeks bindt aan het CARD-motief van deze eiwitten en hun oligomerisatie induceert20. Activering van caspase-4 of caspase-5 bevordert de afgifte van IL-1β door het induceren van een inflammatoire vorm van celdood die pyroptose wordt genoemd door splitsing van het porievormende eiwit gasdermin D (GSDMD)18,19. Bovendien induceert de efflux van kaliumionen als gevolg van caspase-4 en GSDMD-gemedieerde pyroptotische dood activering van het NLRP3-inflammasoom en daaropvolgende activering van caspase-1 22,23. Daarom worden caspase-4, -5 en -11 beschouwd als intracellulaire sensoren voor LPS die pyroptose en caspase-1-activering kunnen induceren als reactie op specifieke stimuli11,24.
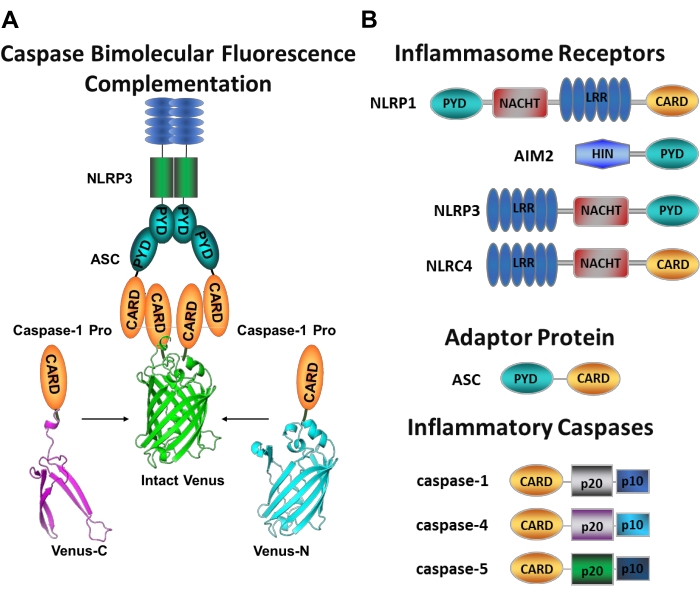
Figuur 1: Inflammatoire caspasen en caspase-bimoleculaire fluorescentiecomplementatie (BiFC) assay. (A) Diagram met het caspase-BiFC-systeem, waarbij twee caspase-1 prodomeinen (C1-pro) gekoppeld aan elk niet-fluorescerend fragment van Venus (Venus-C of Venus-N) worden gerekruteerd naar het NLRP3-activeringsplatform, waardoor Venus wordt gedwongen om opnieuw te vouwen en te fluoresceren. Dit complex verschijnt als een groene vlek onder de microscoop en dient als een uitlezing voor inflammatoire caspase-geïnduceerde nabijheid, wat de eerste stap is in initiator caspase-activering. (B) Schema dat de domeinorganisatie van inflammasoomcomponenten en inflammatoire caspasen weergeeft. Klik hier om een grotere versie van deze figuur te bekijken.
Het meten van specifieke initiator caspases activering is moeilijk, en er zijn niet veel methoden beschikbaar om dit te doen door middel van imaging benaderingen. Caspase Bimolecular Fluorescence Complementation (BiFC) kan worden gebruikt om inflammatoire caspase-activering direct in levende cellen te visualiseren (figuur 1A)25. Deze techniek is onlangs aangepast voor gebruik in menselijke monocyten-afgeleide macrofagen (MDM)21. Caspase BiFC meet de eerste stap in inflammatoire caspase-activering, geïnduceerde nabijheid om dimerisatie te vergemakkelijken. Expressie van plasmiden die coderen voor het CARD-bevattende caspase-prodomein gefuseerd tot niet-fluorescerende fragmenten van het fotostabiele gele fluorescerende eiwit Venus (Venus-C [VC]) en Venus-N [VN]) worden gebruikt. Wanneer de twee caspase-prodomeinen worden gerekruteerd naar hun activeringsplatform of geïnduceerde nabijheid ondergaan, worden de twee helften van Venus in de nabijheid gebracht en gedwongen om opnieuw te vouwen en te fluoresceren (zie figuur 1A,B). Dit biedt een real-time uitlezing van specifieke inflammatoire caspase-activering.
Menselijke MDM brengt overvloedig inflammasoomgenen en patroonherkenningsreceptoren tot expressie die gevaarsignalen en pathogene producten identificeren. Dit biedt een ideaal celtype voor de ondervraging van inflammatoire caspase-routes. Bovendien kunnen ze worden afgeleid uit perifeer bloed en zelfs uit patiëntmonsters om inflammatoire caspase-activering in een specifieke ziektetoestand te beoordelen. Dit protocol beschrijft hoe de BiFC caspase-verslaggevers in MDM kunnen worden geïntroduceerd met behulp van nucleofection, een op elektroporatie gebaseerde transfectiemethode, hoe de cellen moeten worden behandeld om inflammatoire caspase-activering te induceren en hoe de actieve caspase-complexen kunnen worden gevisualiseerd met behulp van microscopiebenaderingen. Bovendien kan deze methodologie worden aangepast om de moleculaire samenstelling van deze complexen, subcellulaire lokalisatie, kinetiek en grootte van deze zeer geordende structuren te bepalen 25,26,27.
