Каспазы представляют собой семейство цистеиновых аспартатных протеаз, которые могут быть сгруппированы в инициирующие каспазы и каспазы палача. Каспазы палача включают каспазу-3, -6 и -7. Они естественным образом обнаруживаются в клетках в виде димеров и расщепляются инициаторами каспаз для выполнения апоптоза1. К каспазам-инициаторам относятся человеческие каспазы-1, -2, -4, -5, -8, -9, -10 и -12. Они обнаруживаются в виде неактивных зимогенов (прокаспасов), которые активируются бесконтактно-индуцированной димеризацией и стабилизируются автопротеолитическим расщеплением 2,3. Воспалительные каспазы являются подмножеством инициирующих каспаз2 и охватывают каспазу-1, -4, -5 и -12 у людей и каспазу-1, -11 и -12 у мыши 4,5. Вместо апоптотической роли, они играют центральную роль в воспалении. Они опосредуют протеолитическую обработку и секрецию про-интерлейкина (IL)-1β и про-IL-18 6,7, которые являются первыми цитокинами, высвобождаемыми в ответ на патогенные захватчики 8,9. Caspase-1 активируется при наборе на его платформу активации; большой молекулярно-массовый белковый комплекс, называемый инфламмасомой (рисунок 1А)10. Димеризация каспазы-4, -5 и -11 происходит независимо от этих платформ через неканонический воспалительный путь11,12.
Канонические инфламмасомы представляют собой цитозольные многомерные белковые комплексы, которые состоят из белка датчика воспаления, адапторного белка ASC (апоптоз-ассоциированный спекоподобный белок, содержащий CARD) и эффекторного белка каспазы-110. Наиболее хорошо изученными каноническими инфламмасомами являются NOD-подобное семейство рецепторов, содержащее пириновый домен (NLRP), NLRP1 и NLRP3, семейство NLR, содержащее CARD (NLRC), NLRC4 и отсутствующее в меланоме 2 (AIM2). Каждый из них содержит пириновый домен, CARD или оба домена. Домен CARD опосредует взаимодействие между CARD-содержащими каспазами и их восходящими активаторами. Поэтому молекула каркаса ASC, которая состоит из N-концевого пиринового домена (PYD) и мотива C-концевой CARD 13,14, необходима для рекрутирования каспазы-1 в воспалительные модели NLRP110, NLRP315 и AIM216.
Каждая инфламмасома названа в честь своего уникального сенсорного белка, который распознает различные провоспалительные стимулы (рисунок 1B). Активаторы этого пути называются каноническими стимулами. Инфламмасомы служат датчиками микробных компонентов и тканевого стресса и собираются, чтобы вызвать сильную воспалительную реакцию путем активации воспалительных каспаз17. Сборка инфламмасом инициирует активацию каспазы-1 для опосредования созревания и секреции ее основных субстратов pro-IL-1β и pro-IL-18. Этот процесс происходит с помощью двухэтапного механизма. Во-первых, прайминговый стимул повышает экспрессию определенных белков инфламмасомы и про-IL-1β путем активации пути NF-κB. Во-вторых, внутриклеточный (канонический) стимул индуцирует сборку и набор прокаспазы-1 6,7.
Каспаза-4 и каспаза-5 являются человеческими ортологами мышиной каспазы-1111. Они активируются инфламмасомно-независимым образом внутриклеточным липополисахаридом (ЛПС), молекулой, обнаруженной во внешней мембране грамотрицательныхбактерий 18,19,20, и внеклеточным гемом, продуктом гемолиза эритроцитов21. Было высказано предположение, что ЛПС связывается непосредственно с мотивом CARD этих белков и индуцирует их олигомеризацию20. Активация каспазы-4 или каспазы-5 способствует высвобождению IL-1β путем индуцирования воспалительной формы гибели клеток, называемой пироптозом, путем расщепления порообразующего белка гасдермина D (GSDMD)18,19. Кроме того, отток ионов калия, образующийся в результате каспазы-4 и GSDMD-опосредованной пироптотической смерти, индуцирует активацию воспалительной системы NLRP3 и последующую активацию каспазы-122,23. Поэтому каспаза-4, -5 и -11 считаются внутриклеточными датчиками для ЛПС, которые способны индуцировать пироптоз и активацию каспазы-1 в ответ на специфические стимулы11,24.
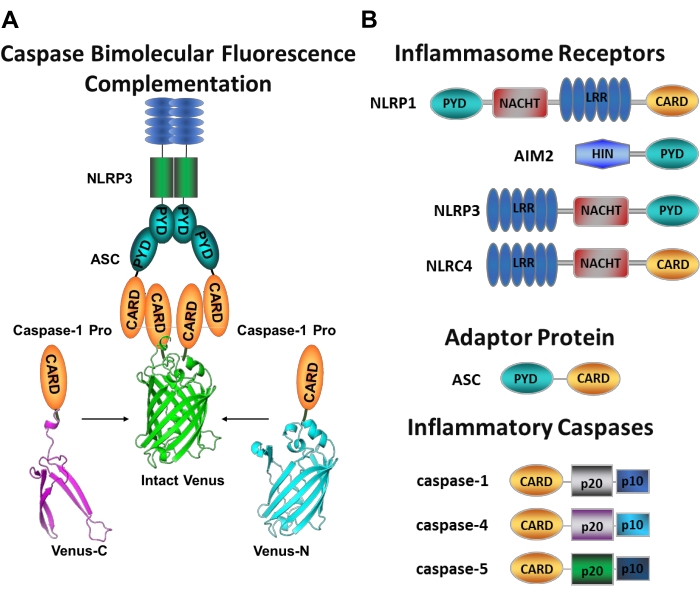
Рисунок 1: Анализ воспалительных каспаз и каспазо-бимолекулярной флуоресцентной комплементации (BiFC). (A) Диаграмма, показывающая систему каспазы-BiFC, где два продомена каспазы-1 (C1-pro), связанные с каждым нефлуоресцентным фрагментом Венеры (Venus-C или Venus-N), набираются на платформу активации NLRP3, заставляя Венеру пересворачиваться и флуоресцентировать. Этот комплекс появляется в виде зеленого пятна под микроскопом и служит считыванием для воспалительной близости, вызванной каспазой, что является первым шагом в активации инициатора каспазы. (B) Схема, показывающая доменную организацию компонентов воспаления и воспалительных каспаз. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Измерение активации конкретных инициирующих каспаз затруднено, и существует не так много методов, доступных для этого с помощью подходов к визуализации. Caspase Bimolecular Fluorescence Complementation (BiFC) может быть использована для визуализации воспалительной активации каспазы непосредственно в живых клетках (Рисунок 1A)25. Этот метод был недавно адаптирован для использования в макрофагах человека, полученных из моноцитов (MDM)21. Caspase BiFC измеряет первый шаг в воспалительной активации каспазы, индуцированной близостью для облегчения димеризации. Использована экспрессия плазмид, кодирующих CARD-содержащий каспазный продомен, слитый с нефлуоресцентными фрагментами фотостабильного желтого флуоресцентного белка Venus (Venus-C [VC]) и Venus-N [VN]). Когда два продомена каспазы набираются на их платформу активации или подвергаются индуцированной близости, две половины Венеры приближаются в непосредственной близости и вынуждены переворачиваться и флуоресцентировать (см. Рисунок 1A, B). Это обеспечивает считывание в режиме реального времени специфической активации воспалительной каспазы.
Человеческий МДМ обильно экспрессирует гены воспаления и рецепторы распознавания образов, которые идентифицируют сигналы опасности и патогенные продукты. Это обеспечивает идеальный тип клеток для опроса воспалительных путей каспазы. Кроме того, они могут быть получены из периферической крови и даже из образцов пациентов для оценки активации воспалительной каспазы в определенном болезненном состоянии. Этот протокол описывает, как вводить репортеры каспазы BiFC в MDM с использованием нуклеофекции, метода трансфекции на основе электропорации, как обрабатывать клетки для индуцирования активации воспалительной каспазы и как визуализировать активные комплексы каспазы с использованием подходов микроскопии. Кроме того, эта методология может быть адаптирована для определения молекулярного состава этих комплексов, субклеточной локализации, кинетики и размера этих высокоупорядоченных структур 25,26,27.
