La maggior parte delle proteine eucariotiche complesse subisce elaborate modificazioni post-traduzionali dopo l’espressione, che richiedono un ripiegamento proteico altamente assistito e co-fattori per essere funzionali1. La produzione di grandi quantità di proteine umane solubili in un ospite batterico rimane una sfida significativa a causa dei costi elevati e della mancanza di solidi metodi di espressione e purificazione, anche per esperimenti di laboratorio su scala ridotta2,3. Le MMP, endopeptidasi umane con grande peso molecolare, sono solitamente espresse come corpi di inclusione insolubili quando espresse in E. coli. L’estrazione di MMP umane solubili porta spesso a un laborioso e dispendioso processo di solubilizzazione e ripiegamento4.
Le MMP hanno ruoli critici sia nei processi fisiologici che in quelli patogenetici. Le MMP umane sono una famiglia di 23 endopeptidasi di zinco, classificate per struttura e specificità del substrato, ed espresse in modo differenziale nonostante un dominio catalitico altamente conservato5,6. Le MMP sono secrete come zimogeni inattivi, regolati tramite attivazione post-traduzionale e loro inibitori endogeni, inibitori tissutali delle metalloproteinasi (TIMP)7,8,9,10. Sebbene inizialmente riconosciute per il loro ruolo nel turnover ECM, le MMP sono state anche implicate nello sviluppo, nella morfogenesi, nella riparazione dei tessuti e nel rimodellamento8. La disregolazione delle MMP è stata in particolare collegata al cancro insieme a malattie neurodegenerative, cardiovascolari e fibrotiche, tra le altre malattie5,7.
Lo sviluppo di solidi metodi di produzione MMP su larga scala è fondamentale per garantire il successo di studi futuri sui meccanismi MMP attraverso saggi biochimici e cellulari. Varie MMP sono state precedentemente espresse in batteri11, comprese le MMP con tag Hisx6, senza alterare l’attività MMP12,13,14,15. Tuttavia, questi metodi includono passaggi noiosi e lunghi che potrebbero essere difficili da replicare.
Le cellule di mammifero possono anche essere utilizzate per esprimere molte proteine umane diverse, garantendo al contempo le corrette modifiche post-traduzionali16. Sebbene il sistema di espressione dei mammiferi sia la scelta ideale per produrre proteine umane ricombinanti con adeguate modifiche post-traduzionali, i principali svantaggi di questo metodo sono le basse rese iniziali, i costosi mezzi di crescita e reagenti, le lunghe tempistiche per raggiungere linee di espressione stabili e il rischio di contaminazione con altre specie come funghi o batteri2,11 . Inoltre, la produzione di MMP nelle linee cellulari di mammiferi produce impurità da proteine cellulari associate come TIMP o fibronectine11. A differenza della lenta crescita cellulare osservata nelle cellule di mammifero, il sistema di espressione batterica offre una produzione proteica su larga scala in un breve periodo insieme a mezzi e requisiti di crescita più semplici. Tuttavia, a causa della mancanza di altre proteine cellulari associate (ad esempio, TIMP) nei sistemi di espressione batterica, le MMP attive a concentrazioni più elevate sono soggette a degradazione attraverso l’autoproteolisi, con conseguente scarsa resa MMP17.
Questo documento descrive un metodo dettagliato per l’espressione batterica, la purificazione e l’attivazione di Hisx6-pro-MMP-3cd ricombinante utilizzando E. coli come ospite di espressione grazie alla sua convenienza, semplicità e successo nel produrre rese più elevate di MMP2,3,18. Poiché E. coli non ha il meccanismo di ripiegamento delle proteine e l’elaborazione post-traduzionale necessari per le MMP ricombinanti e altre proteine complesse, molti ceppi di E. coli sono stati progettati per superare queste limitazioni, rendendo E. coli un ospite più adatto per l’espressione di MMP-3cd umano ricombinante,19,20 . Ad esempio, il ceppo R2DP utilizzato in questo studio migliora l’espressione eucariotica fornendo un plasmide resistente al cloramfenicolo contenente codoni raramente usati in E. coli.
Come descritto in questo protocollo, dopo la sovraespressione di corpi di inclusione relativamente puri dal vettore pET-3a (Figura 1) nelle cellule R2DP, le proteine del dominio catalitico Hisx6-pro-MMP-3 (MMP-3cd) vengono estratte e denaturate4. Hisx6-pro-MMP-3cd3,19 è stato purificato utilizzando la cromatografia con tag di affinità. Dopo il ripiegamento e la dialisi, il pro-MMP-3cd (zimogeno) è stato attivato dall’acetato di 4-aminofenilmercurio (APMA) e l’analisi SDS-PAGE viene utilizzata per valutare le rese e la necessità di un’ulteriore purificazione5,21. Questo protocollo descrive l’espressione, la purificazione e l’attivazione di MMP-3cd solubile come esempio. Tuttavia, può anche essere usato come guida per l’espressione di altre MMP e proteasi umane con espressione simile e meccanismi di attivazione (Figura 2). Per altre proteine diverse da MMP-3cd, si consiglia al lettore di determinare le composizioni tampone e i metodi ottimali per la loro proteina bersaglio prima di tentare questo protocollo.
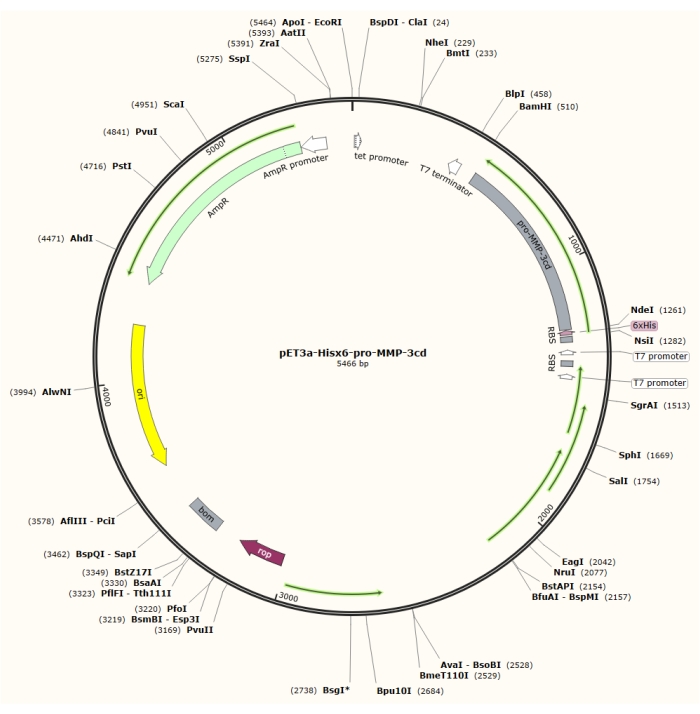
Figura 1: Mappa plasmidica del plasmide pET-3a-Hisx6-pro-MMP-3cd. Il vettore pET-3a include un gene di resistenza all’ampicillina. Una sequenza di tag Hisx6 N-terminale viene clonata nel vettore basato su pET-3a, incluso pro-MMP-3cd, per produrre il costrutto pET-3a-Hisx6-pro-MMP-3cd sotto il controllo del promotore T7 tra i siti di restrizione BamHI e NdeI. Fare clic qui per visualizzare una versione più grande di questa figura.
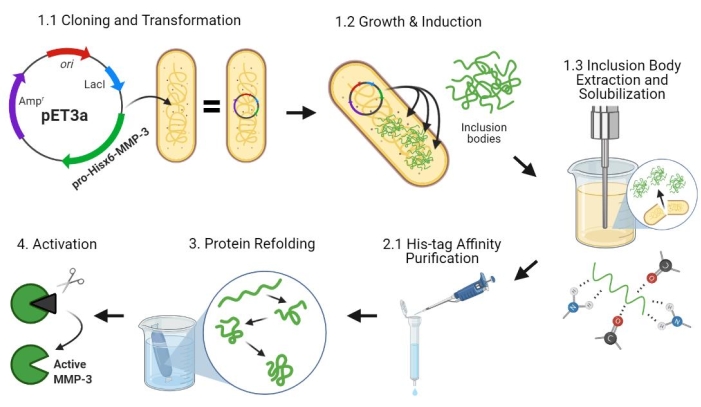
Figura 2: Espressione batterica del plasmide pro-MMP-3cd, purificazione, ripiegamento e attivazione. 1.1: il plasmide pET-3a-Hisx6-pro-MMP-3cd è stato trasformato in cellule BL21(DE3) o R2DP. 1.2: L’espressione proteica Pro-MMP-3cd è stata indotta utilizzando IPTG. 1.3: La lisi chimica e la sonicazione vengono utilizzate per estrarre le proteine Hisx6-pro-MMP-3cd che sono principalmente insolubili e si trovano nei corpi di inclusione. L’urea è stata utilizzata per denaturare e solubilizzare le proteine dai corpi di inclusione. 2.1. La proteina Hisx6-pro-MMP-3cd denaturata è stata purificata tramite purificazione cromatografica di affinità. 3. L’Hisx6-pro-MMP-3cd eluito è stato lentamente ripiegato durante la dialisi attraverso la graduale rimozione dell’urea dal tampone. 4. Infine, la proteina MMP-3cd ripiegata è stata attivata utilizzando APMA rimuovendo il dominio pro-peptide N-terminale. L’APMA viene successivamente rimosso dalla soluzione attraverso la desalinizzazione. I numeri corrispondono alle sezioni di protocollo che descrivono questi passaggi. Abbreviazioni: MMP-3cd = Matrix metalloproteinase-3 catalytic domain; APMA = acetato di 4-aminofenilmercurio. Fare clic qui per visualizzare una versione più grande di questa figura.
