光応答性タンパク質による生物学的プロセスの制御であるオプトジェネティクスは、化学的およびタンパク質生産のための微生物発酵を動的に制御する新しい戦略を提供します1,2。操作された代謝経路の負担と、一部の中間体および生成物の毒性は、しばしば細胞増殖を損なう3。このようなストレスは、バイオマスの蓄積不良や生産性の低下につながる可能性があります3。この課題は、発酵を時間的に成長段階と生産段階に分け、代謝資源をバイオマス蓄積または製品合成にそれぞれ捧げることによって対処することができます4。我々は最近、この二相発酵における成長から生産への移行が、照明条件の変化によって誘発され得ることを示した5,6,7。光入力の高い同調性、可逆性、直交性8は、従来の二相発酵の動的制御に使用される化学誘導物質では複製が困難または不可能な光制御発酵に独自の利点を提供します4,9,10,11。
エリスロバクター・リトラリス由来の青色光応答性EL222タンパク質は、サッカロミセス・セレビシエ5,7,12,13における代謝工学のためのいくつかの光遺伝学的回路を開発するために使用されている。EL222は、青色光活性化(465nm)時に立体構造シフトを受ける光酸素電圧センサ(LOV)ドメインを含み、これにより、同族DNA配列(C120)13に結合することができる。EL222をウイルスVP16活性化ドメイン(VP16-EL222)に融合させることで、合成プロモーターPC120からS. cerevisiae7および他の生物14における遺伝子発現を可逆的に活性化することができる青色光応答性転写因子が得られる。EL222に基づくいくつかの回路が開発され、目的の遺伝子がPC120から直接発現される基本的な光活性化OptoEXPシステム7など、S. cerevisiaeの化学生産に使用されています(図1A)。しかし、発酵の生産段階で典型的に遭遇する高い細胞密度での光の浸透の懸念は、OptoINVRTおよびOptoQ-INVRT回路(図1B)5、7、13のような暗闇で誘導される反転回路を開発する動機となった。これらの系は、それぞれセレビシエ菌およびクラッサ菌由来のガラクトース(GAL)またはキナ酸(Q)レギュロンを利用し、対応するリプレッサー(GAL80およびQS)をVP16-EL222で制御して、光では遺伝子発現を抑制し、暗闇では強く誘導します。OptoEXP回路とOptoINVRT回路を組み合わせることで、遺伝子発現の双方向制御が可能になり、成長期が青色光で誘導され、生産期が暗さで誘導される2相発酵が可能になります(図2A)5,7。
生産段階で遺伝子発現を誘導するために暗闇の代わりに光を使用することは、光遺伝学的制御の能力を大幅に拡大するが、発酵のこの段階で典型的に遭遇する高い細胞密度の光浸透制限を克服することも必要である。この目的のために、我々は、青色光刺激に対する転写応答を増幅するOptoAMPおよびOptoQ-AMPとして知られる回路を開発しました。これらの回路は、VP16-EL222の野生型または過敏性変異体を使用して、GALまたはQレギュロンの転写活性化剤Gal4pまたはQF2の産生をそれぞれ制御し、光による感度の向上とより強い遺伝子発現を達成します12,13(図1C)。OptoAMP回路は、光学密度(600nmで測定)で5Lバイオリアクターで完全かつ均質な光誘導を達成することができます。OD600)は、照明のわずか〜0.35%(バルク表面の〜7%のみに5%の光線量)を有する少なくとも40の値である。これは、100%に近い照明を必要とするOptoEXPと比較して、より高い感度を示しています12。高い細胞密度で光で遺伝子発現を効果的に誘導する能力は、発酵の動的制御のための新しい機会を開きます。これには、化学生産を最適化するために独自の光スケジュールで成長、誘導、生産段階が確立される三相発酵など、2つ以上の時間的段階における発酵の操作が含まれます(図2B)12。
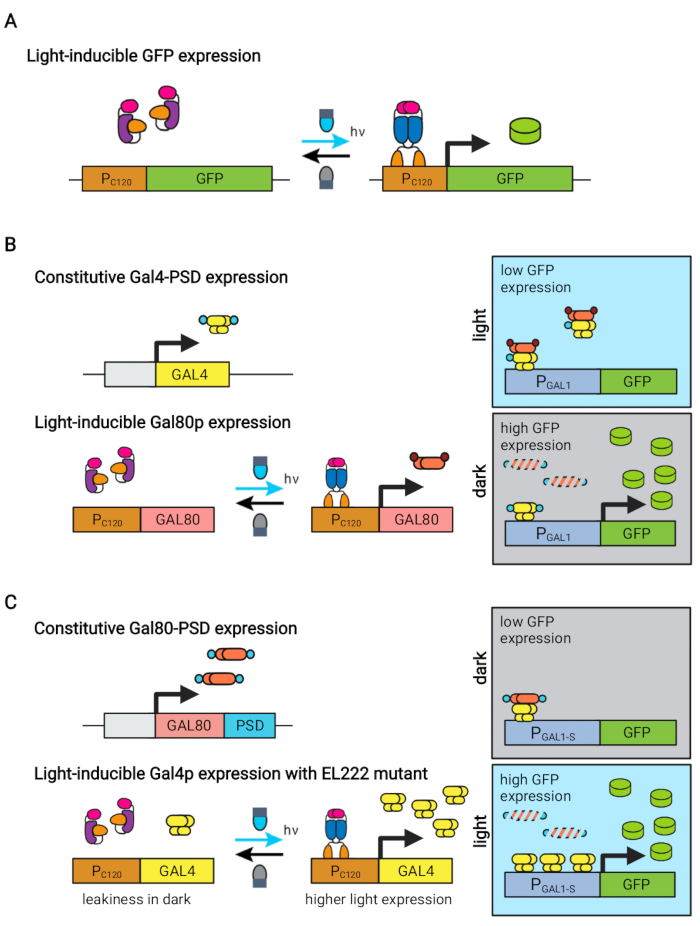
図1:セレビシエ菌の動的制御のための光遺伝学的回路 OptoEXP、OptoINVRT、およびOptoAMP回路は、光に敏感なVP16-EL222システムに基づいています。(A)OptoEXP回路では、青色光への曝露はVP16-EL222の立体構造変化および二量体化を引き起こし、DNA結合ドメインを露出させ、PC120からの転写を可能にする。この図はZhao et al.7から修正されたものである。(B)OptoINVRT回路は、GALまたはQレギュロンを利用して、暗闇での発現を誘導する。GALベースの回路では、VP16-EL222およびGAL4が恒常的に発現され、PC120はGAL80リプレッサーの発現を駆動する(Qベースの回路では、GAL4およびGAL80はそれぞれQF2およびQSに置き換えられ、GALプロモーターの代わりに合成QUAS含有プロモーターが使用される)。光の中で、Gal80pはPGAL1からの目的遺伝子の活性化を妨げる。暗闇では、GAL80は発現されず、構成的デグロンドメイン(小さな茶色のドメイン)に融合することによって急速に分解され、Gal4pによるPGAL1の活性化を可能にする。この図はZhao et al.5から修正されています。(C) OptoAMP回路はVP16-EL222を使用してGALまたはQレギュロンも制御します。これらの回路では、GAL80リプレッサー(またはQS)が恒常的に表現され、光感受性デグロン(小さな青色領域)に融合され、暗闇でのタイトな抑制を保証します。PC120および過敏性VP16−EL222変異体は、光でGAL4(またはQF2)の発現を制御し、光中でPGAL1(またはQUAS含有プロモーター)を強く活性化する。GAL由来の回路は、活性が増大したPGAL1-MやPGAL1-Sなどの操作型PGAL1や、GALレギュロン(PGAL1、PGAL10、PGAL2、PGAL7)によって制御される野生型プロモーターを使用することができる。この図はZhao et al.12から修正されています。この図の拡大版を見るにはここをクリックしてください。
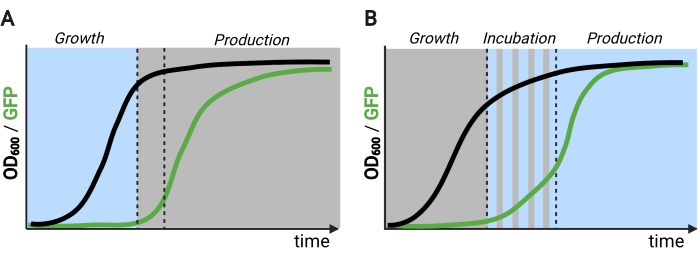
図2:時間の経過に伴う 二相発酵と三相発酵。(A)逆回路で操作される二相発酵は、光駆動の成長段階と暗黒生産段階からなる。成長段階では、生産経路が抑制されたままであるため、バイオマスは蓄積します。所望のOD600に達すると、細胞は、産生段階のために新鮮な培地に再懸濁される前に代謝的に調整するために暗所にシフトされる。(B)三相プロセスにおいて、成長、インキュベーション、および生産段階は、暗い成長期間、パルスインキュベーション、および完全に照らされた生産段階からなることができる固有の光スケジュールによって定義される。バイオレンダーで作成した図。 この図の拡大版を表示するには、ここをクリックしてください。
光遺伝学的回路は、大腸菌における化学的およびタンパク質産生の動的制御のためにも開発されている。OptoLAC回路は、YF1/FixJ 2成分系をベースとする光応答性pDawn回路を使用して、細菌のLacIリプレッサを制御します6(図3)。OptoINVRT5と同様に、OptoLAC回路は、光の中で遺伝子発現を抑制し、暗闇の中でそれを誘導するように設計されています。OptoLAC回路を用いた発現レベルは、標準的なイソプロピルβ-d-1-チオガラクトピラノシド(IPTG)誘導で達成された発現レベルに匹敵するか、それを超えることができるため、化学的誘導の強度を維持しながら、同調性と可逆性を高めます6。したがって、OptoLAC回路は、大腸菌における代謝工学のための効果的な光遺伝学的制御を可能にする。
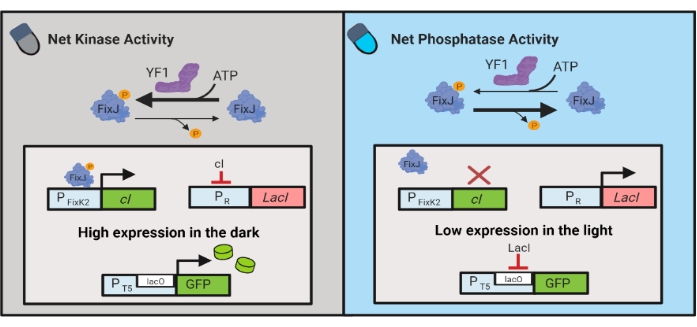
図3:大腸菌の動的制御のためのOptoLAC回路。 OptoLAC回路は、pDawnシステムとlacオペロンを適応させて、暗闇の中での活性化と光での抑圧を実現します。暗所では、YF1はFixJをリン酸化し、PFixK2プロモーターを活性化してcIリプレッサーを発現させる。cIリプレッサーは、PRプロモーターからのlacIリプレッサーの発現を防止し、lacO含有プロモーターからの目的遺伝子の転写を可能にする。逆に、青色光はYF1ネットキナーゼ活性を低下させ、FixJリン酸化を逆転させ、したがってcI発現を逆転させ、lacIの発現を抑制し、lacO含有プロモーターからの発現を妨げる。この図はLalwani et al.6から修正されたものである。この図の拡大版を表示するには、ここをクリックしてください。
ここでは、化学またはタンパク質生産のための セレビシエ 菌および 大腸菌 の光制御発酵のための基本的なプロトコルについて説明します。酵母とバクテリアの両方について、まず、光駆動の増殖段階と、OptoINVRTおよびOptoLAC回路によって可能になる暗闇誘発生産段階を有する発酵に焦点を当てています。続いて、OptoAMP回路によって可能になる三相(成長、誘導、生産)光制御発酵のプロトコルについて説明します。さらに、光遺伝学的に制御された発酵をマイクロプレートからラボ規模のバイオリアクターにスケールアップする方法についても説明します。このプロトコルにより、化学的またはタンパク質生産のための光制御発酵を実行するための完全で再現性の高いガイドを提供することを目指しています。
