La optogenética, el control de procesos biológicos con proteínas sensibles a la luz, ofrece una nueva estrategia para controlar dinámicamente las fermentaciones microbianas para la producción química y proteica1,2. La carga de las vías metabólicas modificadas y la toxicidad de algunos productos intermedios y productos a menudo perjudican el crecimiento celular3. Tales tensiones pueden conducir a una acumulación deficiente de biomasa y a una reducción de la productividad3. Este desafío puede abordarse dividiendo temporalmente las fermentaciones en una fase de crecimiento y producción, que dedican recursos metabólicos a la acumulación de biomasa o a la síntesis de productos, respectivamente4. Recientemente demostramos que la transición del crecimiento a la producción en esta fermentación bifásica puede ser inducida con cambios en las condiciones de iluminación5,6,7. La alta capacidad de ajuste, reversibilidad y ortogonalidad de las entradas de luz8 ofrece ventajas únicas a las fermentaciones controladas por la luz que son difíciles o imposibles de replicar con inductores químicos utilizados en el control dinámico de fermentaciones bifásicas convencionales4,9,10,11.
La proteína EL222 sensible a la luz azul derivada de Erythrobacter litoralis se ha utilizado para desarrollar varios circuitos optogenéticos para la ingeniería metabólica en Saccharomyces cerevisiae5,7,12,13. EL222 contiene un dominio sensor de voltaje de luz-oxígeno (LOV) que sufre un cambio conformacional tras la activación de la luz azul (465 nm), lo que le permite unirse a su secuencia de ADN cognado (C120)13. La fusión de EL222 al dominio de activación viral VP16 (VP16-EL222) da como resultado un factor de transcripción sensible a la luz azul que puede activar reversiblemente la expresión génica en S. cerevisiae7 y otros organismos14 a partir del promotor sintético PC120. Varios circuitos basados en EL222 han sido desarrollados y utilizados para la producción química en S. cerevisiae, como el sistema básico OptoEXP activado por luz7, en el que el gen de interés se expresa directamente desde PC120 (Figura 1A). Sin embargo, las preocupaciones de penetración de la luz en las altas densidades celulares típicamente encontradas en la fase de producción de las fermentaciones nos motivaron a desarrollar circuitos invertidos que se inducen en la oscuridad, como los circuitos OptoINVRT y OptoQ-INVRT (Figura 1B)5,7,13. Estos sistemas aprovechan los reguleones de galactosa (GAL) o ácido quínico (Q) de S. cerevisiae y N. crassa, respectivamente, controlando sus correspondientes represores (GAL80 y QS) con VP16-EL222, para reprimir la expresión génica a la luz e inducirla fuertemente en la oscuridad. La combinación de los circuitos OptoEXP y OptoINVRT da como resultado un control bidireccional de la expresión génica, permitiendo fermentaciones bifásicas en las que la fase de crecimiento se induce con luz azul y la fase de producción con oscuridad (Figura 2A)5,7.
El uso de la luz en lugar de la oscuridad para inducir la expresión génica durante la fase de producción ampliaría en gran medida las capacidades de los controles optogenéticos, pero también requeriría superar las limitaciones de penetración de la luz de las altas densidades celulares que se encuentran típicamente en esta fase de fermentación. Para ello, hemos desarrollado circuitos, conocidos como OptoAMP y OptoQ-AMP, que amplifican la respuesta transcripcional a la estimulación de la luz azul. Estos circuitos utilizan mutantes de tipo salvaje o hipersensibles de VP16-EL222 para controlar la producción de los activadores transcripcionales Gal4p o QF2 de los reguleones GAL o Q, respectivamente, logrando una mayor sensibilidad y una expresión génica más fuerte con luz12,13 (Figura 1C). Los circuitos OptoAMP pueden lograr una inducción de luz completa y homogénea en biorreactores de 5 L a una densidad óptica (medida a 600 nm; OD600) valores de al menos 40 con solo ~ 0.35% de iluminación (dosis de luz del 5% en solo ~ 7% de la superficie a granel). Esto demuestra un mayor grado de sensibilidad en comparación con OptoEXP, que requiere cerca del 100% de iluminación12. La capacidad de inducir eficazmente la expresión génica con luz a altas densidades celulares abre nuevas oportunidades para el control dinámico de las fermentaciones. Esto incluye fermentaciones operativas en más de dos fases temporales, como las fermentaciones trifásicas, en las que se establecen fases de crecimiento, inducción y producción con programas de luz únicos para optimizar la producción química (Figura 2B)12.
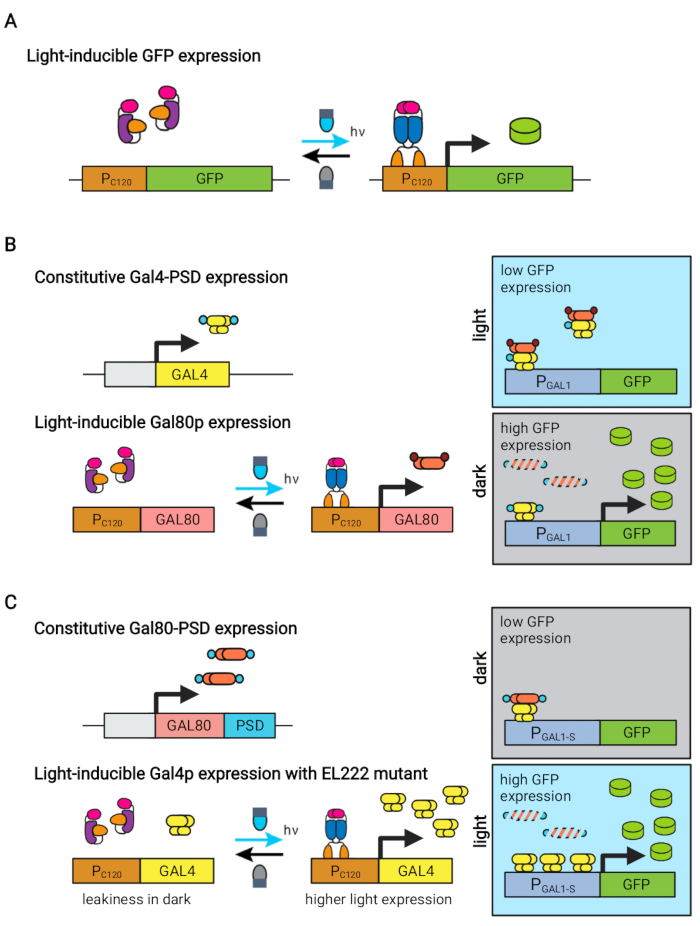
Figura 1: Circuitos optogenéticos para el control dinámico de S. cerevisiae. Los circuitos OptoEXP, OptoINVRT y OptoAMP se basan en el sistema VP16-EL222 sensible a la luz. (A) En el circuito OptoEXP, la exposición a la luz azul causa un cambio conformacional y dimerización de VP16-EL222, que expone un dominio de unión al ADN y permite la transcripción desde PC120. La cifra ha sido modificada a partir de Zhao et al.7. (B) Los circuitos OptoINVRT aprovechan los regulones GAL (mostrados) o Q para inducir la expresión en la oscuridad. En los circuitos basados en GAL, VP16-EL222 y GAL4 se expresan constitutivamente, mientras que PC120 impulsa la expresión del represor GAL80 (en los circuitos basados en Q, GAL4 y GAL80 se reemplazan por QF2 y QS, respectivamente, y se utiliza un promotor sintético que contiene QUAS en lugar de un promotor GAL). A la luz, Gal80p previene la activación del gen de interés de PGAL1. En la oscuridad, GAL80 no se expresa y se degrada rápidamente fusionándolo a un dominio degron constitutivo (pequeño dominio marrón), que permite la activación de PGAL1 por Gal4p. La cifra ha sido modificada a partir de Zhao et al.5. (C) Los circuitos OptoAMP también utilizan VP16-EL222 para controlar los reguleones GAL (mostrados) o Q. En estos circuitos, el represor GAL80 (o QS) se expresa constitutivamente y se fusiona con un degron fotosensible (pequeño dominio azul) que garantiza una represión estricta en la oscuridad. PC120 y una expresión de control mutante hipersensible VP16-EL222 de GAL4 (o QF2) con luz, que activa fuertemente PGAL1 (o un promotor que contiene QUAS) en la luz. Los circuitos derivados de GAL pueden utilizar formas de ingeniería de PGAL1, como PGAL1-M o PGAL1-S, que tienen una mayor actividad, así como promotores de tipo salvaje controlados por el regulón GAL (PGAL1, PGAL10, PGAL2, PGAL7). La figura ha sido modificada de Zhao et al.12. Haga clic aquí para ver una versión más grande de esta figura.
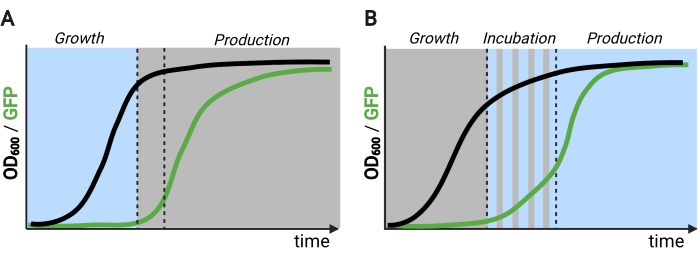
Figura 2: Fermentaciones bifásicas y trifásicas a través del tiempo. (A) Las fermentaciones bifásicas operadas con circuitos invertidos consisten en una fase de crecimiento impulsada por la luz y una fase de producción oscura. En la fase de crecimiento, la biomasa se acumula a medida que la vía de producción permanece reprimida. Al alcanzar el OD600 deseado, las células se desplazan a la oscuridad para ajustarse metabólicamente antes de ser resuspendidas en medios frescos para la fase de producción. (B) En un proceso de tres fases, las fases de crecimiento, incubación y producción están definidas por programas de luz únicos, que pueden consistir en un período de crecimiento oscuro, incubación pulsada y fase de producción completamente iluminada. Figura creada con Biorender. Haga clic aquí para ver una versión más grande de esta figura.
También se han desarrollado circuitos optogenéticos para el control dinámico de la producción química y proteica en E. coli. Los circuitos OptoLAC controlan el represor bacteriano LacI utilizando el circuito pDawn sensible a la luz, que se basa en el sistema de dos componentes YF1/FixJ6 (Figura 3). Al igual que OptoINVRT5, los circuitos OptoLAC están diseñados para reprimir la expresión génica en la luz e inducirla en la oscuridad. Los niveles de expresión que utilizan circuitos OptoLAC pueden igualar o superar los alcanzados con la inducción estándar de isopropilo β-d-1-tiogalactopiranósido (IPTG), manteniendo así la fuerza de la inducción química al tiempo que ofrece una mayor capacidad de ajuste y reversibilidad6. Por lo tanto, los circuitos OptoLAC permiten un control optogenético efectivo para la ingeniería metabólica en E. coli.
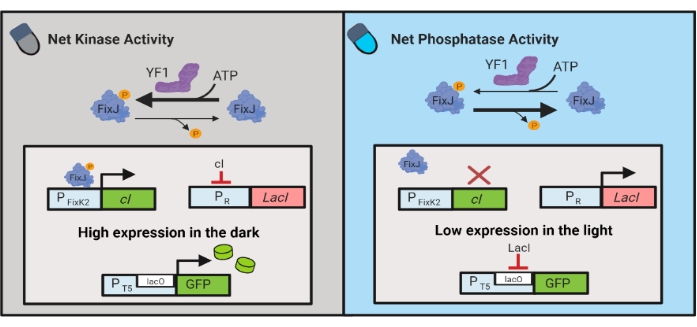
Figura 3: Circuitos OptoLAC para el control dinámico de E. coli. Los circuitos OptoLAC adaptan el sistema pDawn y el operón lac para lograr la activación en la oscuridad y la represión en la luz. En la oscuridad, YF1 fosforila FixJ, que luego activa el promotor PFixK2 para expresar el represor cI . El represor cI impide la expresión del represor lacI del promotor PR , lo que permite la transcripción del gen de interés a partir de un promotor que contiene lacO. Por el contrario, la luz azul reduce la actividad de la quinasa neta YF1, revirtiendo la fosforilación de FixJ y, por lo tanto, la expresión de cI , lo que desreprime la expresión de lacI e impide la expresión del promotor que contiene lacO. La figura ha sido modificada a partir de Lalwani et al.6. Haga clic aquí para ver una versión más grande de esta figura.
Describimos aquí los protocolos básicos para fermentaciones controladas por la luz de S. cerevisiae y E. coli para la producción química o proteica. Tanto para levaduras como para bacterias, primero nos centramos en fermentaciones con una fase de crecimiento impulsada por la luz y una fase de producción inducida por la oscuridad habilitada por los circuitos OptoINVRT y OptoLAC. Posteriormente, describimos un protocolo para una fermentación trifásica (crecimiento, inducción, producción) controlada por la luz habilitada por los circuitos OptoAMP. Además, describimos cómo escalar fermentaciones controladas optogenéticamente desde microplacas hasta biorreactores a escala de laboratorio. Con este protocolo, nuestro objetivo es proporcionar una guía completa y fácilmente reproducible para la realización de fermentaciones controladas por la luz para la producción química o proteica.
