Il metabolismo è il processo cellulare di produzione di energia. Il metabolismo cellulare comprende più vie, tra cui glicolisi, fosforilazione ossidativa e glutaminolisi. Le cellule sane utilizzano queste vie metaboliche per generare energia per la proliferazione e la funzione, come la produzione di citochine da parte delle cellule immunitarie. Molte malattie, tra cui i disturbi metabolici, il cancro e la neurodegenerazione, sono caratterizzate da un metabolismo cellulare alterato1. Ad esempio, alcuni tipi di cellule tumorali hanno tassi elevati di glicolisi, anche in presenza di ossigeno, per generare molecole per la sintesi di acidi nucleici, proteine e lipidi2,3. Questo fenomeno, noto come effetto Warburg, è un segno distintivo di molti tipi di cancro, tra cui il cancro al seno, il cancro ai polmoni e i glioblastomi4. A causa delle alterazioni del metabolismo cellulare associate alla progressione del cancro, il metabolismo cellulare può essere un biomarcatore surrogato per la risposta ai farmaci5,6. Inoltre, comprendere l’efficacia del farmaco a livello cellulare è fondamentale in quanto l’eterogeneità cellulare può portare a diverse risposte farmacologiche negli individui7,8.
Le tecnologie che identificano e quantificano i cambiamenti nel metabolismo cellulare sono essenziali per gli studi sul cancro e sulla risposta ai farmaci. Le analisi chimiche e proteiche vengono utilizzate per valutare il metabolismo delle cellule o dei tessuti, ma mancano di risoluzione unicellulare e di informazioni spaziali. I saggi basati sul lettore di piastre metaboliche possono misurare il pH e il consumo di ossigeno nel campione nel tempo e la successiva perturbazione metabolica da parte di sostanze chimiche. Il pH può essere utilizzato per calcolare il tasso di acidificazione extracellulare (ECAR), che fornisce una panoramica dell’attività glicolitica delle cellule9. I metodi di imaging di tutto il corpo, tra cui la tomografia a emissione di positroni fluoro-D-glucosio 2-[fluoro-18] (FDG PET) e la spettroscopia di risonanza magnetica (MRS), sono modalità di imaging non invasive utilizzate clinicamente per identificare la recidiva del tumore e l’efficacia del farmaco attraverso misurazioni metaboliche10,11,12,13,14.
FDG-PET visualizza l’assorbimento tissutale di FDG, un analogo del glucosio radiomarcato. L’aumento dell’assorbimento di FDG-PET da parte dei tumori rispetto al tessuto circostante è dovuto all’effetto Warburg12,13. MRS visualizza nuclei comuni di molecole utilizzate per il metabolismo, come 13C e 31P, e può ottenere informazioni dinamiche su come il metabolismo cambia in risposta a stimoli, come l’esercizio fisico o il mangiare14. Sebbene FDG-PET e MRS possano essere utilizzati clinicamente, queste tecnologie mancano della risoluzione spaziale per risolvere l’eterogeneità intratumorale. Allo stesso modo, le misurazioni del consumo di ossigeno vengono effettuate su una popolazione di massa di cellule. L’imaging a autofluorescenza supera l’ostacolo della risoluzione spaziale di queste tecnologie e fornisce un metodo non invasivo per quantificare il metabolismo cellulare.
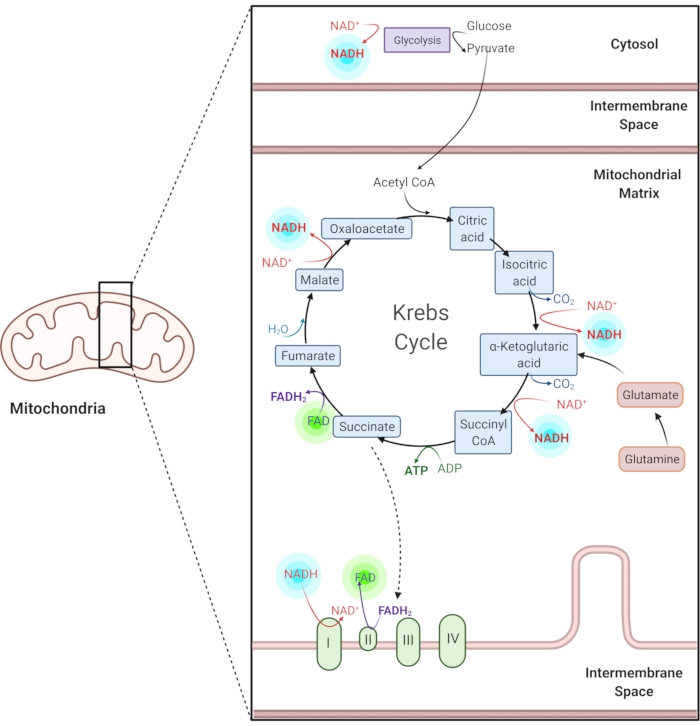
Figura 1: NADH e FAD nelle vie metaboliche comuni. NADH e FAD sono coenzimi utilizzati nella glicolisi, nel ciclo di Krebs e nella catena di trasporto degli elettroni. L’imaging a autofluorescenza di queste molecole fornisce informazioni sul metabolismo cellulare. Fare clic qui per visualizzare una versione più grande di questa figura.
La nicotinamide adenina (fosfato) dinucleotide (NAD(P)H) e la flavina adenina dinucleotide ossidata (FAD) sono coenzimi delle reazioni metaboliche, tra cui glicolisi, fosforilazione ossidativa e glutaminolisi (Figura 1). Sia NAD(P)H che FAD sono autofluorescenti e forniscono un contrasto endogeno per l’imaging a fluorescenza1,15. NADPH ha proprietà fluorescenti simili a NADH. Per questo motivo, NAD(P)H è spesso usato per rappresentare il segnale combinato di NADH e NADPH2,16.
L’imaging a vita di fluorescenza (FLIM) quantifica la durata della fluorescenza o il tempo per il quale un fluoroforo è nello stato eccitato. Le durate di fluorescenza rispondono al microambiente dei fluorofori e forniscono informazioni sul metabolismo cellulare17. NAD(P)H e FAD possono esistere all’interno delle cellule in conformazioni legate a proteine o libere, ognuna delle quali ha una vita diversa. Il NAD(P)H libero ha una vita più breve rispetto al NAD(P)H legato alle proteine; al contrario, la FAD libera ha una durata maggiore rispetto alla FAD18,19 legata. Le durate e i pesi dei componenti a vita possono essere quantificati dai dati di decadimento della durata della fluorescenza tramite Eq. (1)20:
I(t) = α 1e-t/τ1 + α 2e-t/τ2 + C (1)
Eq (1) rappresenta l’intensità di fluorescenza normalizzata in funzione del tempo. Le α 1 e α 2 in questa equazione rappresentano le componenti proporzionali di vita breve e lunga (α 1+ α 2=1), rispettivamente, τ1 e τ2 rappresentano rispettivamente la vita breve e lunga, e C rappresenta la luce di fondo7,20. La durata ponderata in ampiezza, qui rappresentata come τm, è calcolata utilizzando Eq. (2).
τm= α 1τ1+ α 2τ2 (2)
Una vita media può essere calcolata facendo la media di “t” sul decadimento di intensità del fluoroforo, che per un decadimento bieculente è mostrato da Eq. (3)17,21.
τ*m= (α 1τ12+ α 2τ22)/ (α 1τ1+ α 2τ2) (3)
Un’immagine di intensità di fluorescenza può essere calcolata dall’immagine a vita integrando il decadimento della durata della fluorescenza. L’imaging a autofluorescenza è un metodo non distruttivo e privo di etichette che può essere utilizzato per caratterizzare il metabolismo delle cellule vive a una risoluzione subcellulare. Il rapporto redox ottico fornisce una metrica analogica ottica dello stato redox chimico della cellula ed è calcolato come il rapporto tra le intensità NAD(P)H e FAD. Sebbene la formula per il calcolo del rapporto redox ottico non sia standardizzata22,23,24,25, è qui definita come l’intensità di FAD sulle intensità combinate di NAD(P)H e FAD. Questa definizione viene utilizzata perché l’intensità sommata nel denominatore normalizza la metrica tra 0 e 1 e il risultato atteso dell’inibizione del cianuro è una diminuzione del rapporto redox. Le durate di fluorescenza di NAD(P)H e FAD liberi forniscono informazioni sui cambiamenti nel microambiente del solvente metabolico, tra cui pH, temperatura, vicinanza all’ossigeno e osmolarità17.
I cambiamenti nella durata di fluorescenza delle frazioni legate di NAD(P)H e FAD possono indicare l’utilizzo della via metabolica e il metabolismo specifico del substrato26. I pesi dei componenti possono essere interpretati per cambiamenti nella frazione libera al legato dei coenzimi18,19. Complessivamente, queste metriche quantitative di durata dell’autofluorescenza consentono l’analisi del metabolismo cellulare e l’imaging ad autofluorescenza è stato utilizzato per identificare neoplasie da tessuti normali27,28, caratterizzare le cellule staminali29,30, valutare la funzione delle cellule immunitarie31,32,33,34,35, misurare l’attività neurologica36, 37,38 e comprensione dell’efficacia dei farmaci in tipi di cancro come il cancro al seno e il cancro della testa e del collo21,39,40,41,42. L’imaging ad autofluorescenza ad alta risoluzione può essere combinato con la segmentazione dell’immagine per l’analisi di singole cellule e la quantificazione dell’eterogeneità intrapopolazione43,44,45,46,47.
NAD(P)H e FAD possono essere ripresi su microscopi a fluorescenza a fotone singolo o multifotone configurati per l’imaging di intensità o durata. Per i microscopi a singolo fotone, NAD(P)H e FAD sono tipicamente eccitati a lunghezze d’onda di 375-405 nm e 488 nm, rispettivamente, a causa di sorgenti laser comuni a queste lunghezze d’onda48. Nell’eccitazione a fluorescenza a due fotoni, NAD(P)H e FAD ecciteranno a lunghezze d’onda di circa 700-750 nm e da 700 a 900 nm, rispettivamente15,49. Una volta che i fluorofori sono eccitati, NAD(P)H e FAD emettono fotoni a lunghezze d’onda comprese tra ~ 410 nm a ~ 490 nm e ~ 510 nm a ~ 640 nm, rispettivamente15. Le lunghezze d’onda massime di emissione NAD(P)H e FAD sono rispettivamente di circa 450 nm e 535 nm48.
A causa delle loro diverse lunghezze d’onda di eccitazione ed emissione, la fluorescenza dei due coenzimi metabolici può essere isolata spettralmente. Una comprensione delle caratteristiche spettrali di NAD(P)H e FAD è necessaria per la progettazione e l’ottimizzazione dei protocolli di imaging ad autofluorescenza. Il cianuro è un inibitore iv del complesso della catena di trasporto degli elettroni (ETC). Gli effetti del cianuro sul metabolismo cellulare e le intensità di autofluorescenza e la durata di vita di NAD(P)H e FAD all’interno delle cellule sono ben caratterizzati27,40. Pertanto, un esperimento di perturbazione al cianuro è un mezzo efficace per convalidare i protocolli di imaging NAD(P)H e FAD. Un esperimento di cianuro di successo fornisce la certezza che il protocollo di imaging NAD(P)H e FAD può essere utilizzato per valutare il metabolismo di gruppi sconosciuti o perturbazioni.
