Il suolo è un ambiente eccezionalmente diversificato contenente un’abbondanza di microrganismi che sono strumentali ai cicli del carbonio e del fosforo 1,2. I funghi filamentosi sono una componente importante di numerosi ecosistemi come decompositori di materia organica e inorganica e possono migliorare la nutrizione delle piante attraverso la formazione di relazioni simbiotiche 3,4. All’interno del suolo, i funghi interagiscono dinamicamente con una moltitudine di microbi come altri funghi5, batteri6, virus7 e nematodi8. Queste interazioni hanno conseguenze significative per la salute del suolo e delle piante. Tuttavia, a causa della mancanza di sistemi sperimentali appropriati in grado di eseguire l’imaging di microrganismi interagenti ad alta risoluzione, molti rimangono indefiniti.
Le indagini riguardanti le interazioni batterico-fungine (BFI) e le interazioni fungo-fungine (FFI) hanno preziose applicazioni in una vasta gamma di campi, tra cui gli antimicrobici in medicina e gli agenti di controllo biologico in agricoltura. Ad esempio, il fungo Coprinopsis cinerea produce il peptide copsina, che ha dimostrato di esibire attività antibatterica contro il patogeno umano Listeria monocytogenes9. Allo stesso modo, il composto di derivazione fungina, griseofulvina, è ampiamente usato come trattamento per le infezioni fungine umane ed è inoltre in grado di inibire la crescita del fungo patogeno della pianta Alternaria solani10,11. Diversi ceppi del batterio Bacillus subtilis che abita nel suolo hanno anche dimostrato di essere efficaci agenti di biocontrollo del patogeno vegetale fungino Rhizoctonia solani12,13. Tuttavia, a causa delle limitazioni associate alle metodologie tradizionali, i BFI e gli FFA sono poco compresi a livello di singole cellule.
Gli studi convenzionali in genere esplorano BFI e FFI su macroscala usando placche di agar con due o più specie in confronto. La loro interazione è valutata misurando i tassi di crescita e la produzione di metaboliti delle specie in confronto 14,15,16; tuttavia, questa metodologia è risolta solo a livello di colonia. Per studiare le interazioni a livello cellulare, gli inoculanti batterici e fungini possono essere coltivati su vetrini per microscopio rivestiti di agar che vengono poi ripresi al microscopio17. Tuttavia, può essere difficile seguire una singola ifa usando vetrini per microscopio a causa della mancanza di confinamento, il che significa che le immagini time-lapse sono più difficili da ottenere. Inoltre, l’opportunità di confinare spazialmente altri microrganismi all’interno di regioni definite del micelio fungino o di creare ambienti chimici definiti che possono essere perturbati, ad esempio, non è possibile in tali configurazioni. La natura “scatola nera” del suolo aggiunge anche alla complessità dello studio delle interazioni fungo-microbiche a livello di singole cellule18. Osservando le specie interagenti lontano dall’incredibile diversità del microbioma del suolo, è possibile valutare il modo esatto in cui i singoli membri interagiscono. Pertanto, vi è una continua necessità di piattaforme versatili che consentano l’imaging ad alta risoluzione a cella singola di BFI e FFA.
Le tecnologie microfluidiche, i cosiddetti sistemi lab-on-a-chip, forniscono una piattaforma ideale per lo studio di BFI e FONI a livello di singole celle. Il campo della microfluidica, originato da tecnologie sviluppate per l’analisi chimica e la microelettronica, è stato adottato dalle scienze biologiche19. Le tecnologie microfluidiche regolano piccoli volumi di fluidi all’interno di una rete su misura di canali miniaturizzati, aventi almeno una dimensione su scala micrometrica, e il loro uso nella ricerca biologica si sta espandendodi 20. In particolare, sono stati sviluppati dispositivi microfluidici per esaminare la crescita di funghi filamentosi 21,22,23,24,25,26,27,28,29,30. Uno dei vantaggi dell’utilizzo di questa tecnologia è che il confinamento delle ife e la distribuzione dei nutrienti all’interno di microcanali assomiglia più da vicino alla struttura dell’ambiente del suolo rispetto ai metodi convenzionali di agar31. Recentemente, le piattaforme microfluidiche sono state utilizzate per studiare le interazioni tra neutrofili umani e patogeni fungini32, batteri e radici vegetali33, nonché funghi e nematodi34,35.
Uno dei molti vantaggi dell’utilizzo della microfluidica per lo studio delle interazioni microbiche include il controllo specifico dell’ambiente dei microcanali. Ad esempio, i regimi di flusso laminare possono essere sfruttati per generare gradienti di concentrazione definiti, il che è particolarmente utile quando si esamina la chemiotassi batterica36. Un altro vantaggio è che la natura trasparente del poli(dimetilsilossano) (PDMS), un polimero elastomerico economico e biocompatibile comunemente usato nella produzione di dispositivi microfluidici, facilita l’imaging ad alta risoluzione di singole cellule utilizzando la microscopia a campo luminoso e fluorescenza37. Allo stesso modo, il confinamento dei microbi all’interno di microcanali significa che possono essere eseguiti esperimenti time-lapse che tracciano singole cellule, consentendo di registrare e quantificare le singole risposte cellulari37. Infine, poiché i dispositivi microfluidici possono essere progettati per essere facili da usare, possono essere facilmente utilizzati da non esperti38.
Approfondire la conoscenza delle interazioni tra microrganismi che vivono nel suolo è importante per migliorare le pratiche di gestione sostenibile degli ecosistemi che mantengono la biodiversità e per mitigare l’impatto dei cambiamenti climatici sugli ambienti terrestri39. Pertanto, lo sviluppo di nuovi strumenti microfluidici è fondamentale per espandere la comprensione dei funghi e delle loro interazioni a livello cellulare. Il protocollo qui si concentrerà su due dispositivi microfluidici prodotti per lo studio di BFI40 e FFI41 come rappresentato nella Figura 1.
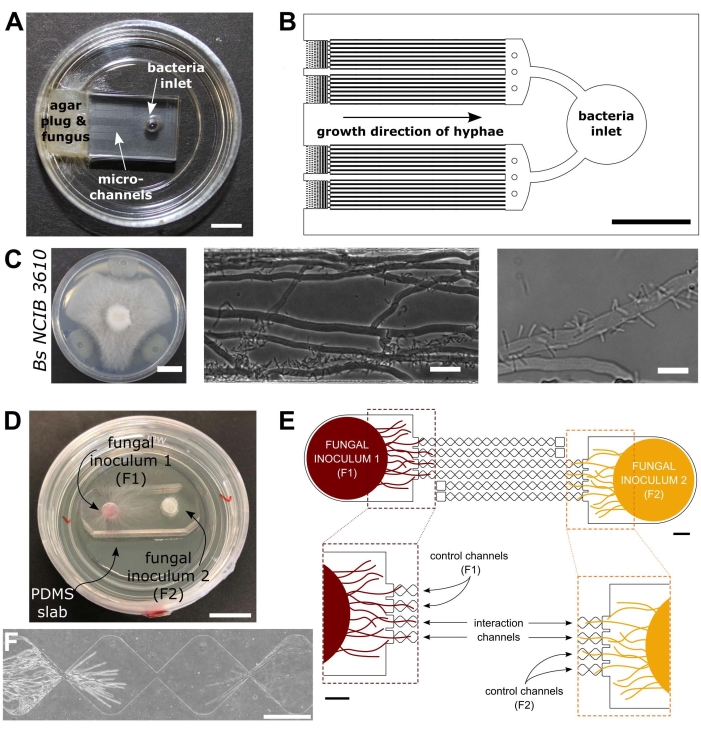
Figura 1: Rappresentazione visiva e schematica dei dispositivi di interazione batterico-fungina (BFI) e di interazione fungo-fungino (FFI). (A) Immagine del dispositivo BFI. Una spina miceliale è posizionata all’ingresso di un’estremità dei microcanali per consentire la crescita ifale nel dispositivo. L’ingresso batterico si trova all’estremità opposta. Barra della scala = 5 mm. (B) Panoramica schematica del dispositivo BFI, raffigurante il posizionamento delle prese batteriche e la direzione della crescita ifale attraverso i microcanali di interazione. I canali sono profondi 10 μm, larghi 100 μm e lunghi 7 mm, con 28 canali di osservazione in totale. (C) Saggio di confronto su piastra di agar tra Coprinopsis cinerea e Bacillus subtilis NCIB 3610, barra di scala = 20 mm (sinistra). Immagini al microscopio che mostrano l’interazione tra C. cinerea e B. subtilis NCIB 3610 all’interno del microcanale (centrale e destro), cioè l’attaccamento polare dei batteri alle ife fungine. Barra della scala = 25 μm (al centro) e 10 μm (a destra). (D) Immagine del dispositivo FFI legato a una capsula di Petri con fondo di vetro, doppia inoculata con tappi miceliali. Barra della scala = 1 cm. (E) Panoramica schematica del dispositivo FFI. Due spine inoculanti fungine vengono introdotte nelle prese d’ingresso alle due estremità del dispositivo, consentendo l’esplorazione ifale dei microcanali. I canali di controllo sono collegati a un solo ingresso fungino e hanno un canale senza uscita, impedendo le interazioni tra i funghi di prova. I canali di interazione collegano entrambe le insenature fungine e consentono interazioni ifali tra i soggetti del test all’interno del microcanale. Ogni canale di interazione è costituito da 18 sezioni a forma di diamante, che misurano una lunghezza totale di 8,8 mm (490 x 430 μm per diamante), 10 μm di profondità e hanno una regione di collegamento tra ciascun diamante di 20 μm. I tipi di canale sono duplicati, barre di scala = 1 mm. (F) Zona di interazione tra due fronti ifali in avvicinamento, che crescono da estremità opposte del canale di interazione interconnesso. Immagine al microscopio a contrasto di fase, barra di scala = 250 μm. I pannelli in questa figura sono stati modificati da Stanley et al., 2014 (A-C)40 e Gimeno et al., 2021 (D-F)41. Fare clic qui per visualizzare una versione più grande di questa figura.
