קולטנים מצומדים לחלבון G (GPCRs), הנמצאים משמרים ועד בני אדם, מייצגים את משפחת-העל הגדולה ביותר של קולטנים באורגניזמים רבים1. הם ממלאים תפקידים קריטיים בוויסות כמעט כל התהליכים הביולוגיים בבעלי חיים. ישנם 50-200 GPCRs בגנום של פרוקי רגליים, כלומר הם מייצגים את משפחת קולטני הממברנההגדולה ביותר 2. הם מסווגים לשש מחלקות עיקריות, A-F, בהתבסס על דמיון הרצף והפונקציות שלהם3. GPCRs מתמרים אותות חוץ-תאיים שונים, כגון אלה של הורמונים, נוירופפטידים, אמינים ביוגניים, גלוטמט, פרוטון, ליפוגליקופרוטאינים ופוטונים4. GPCRs מצמידים לחלבוני G הטרוטרימרים (Gα, Gβ ו-Gγ) כדי להעביר אותות במורד הזרם. GPCRs המצומדים לחלבוני Gαs או Gα i/o מגבירים או מקטינים, בהתאמה, את רמות האדנוזין מונופוספט (cAMP) התוך-תאיים 3′, 5′-מחזורי על-ידיהפעלה או עיכוב של אדניליל ציקלאז. GPCRs המצומדים ל-Gαq/11 גורמים לשחרור סידן ממאגרי הסידן ברשתית האנדופלסמית על-ידי הפעלת מסלול הפוספוליפאז C (PLC)-אינוזיטול-1,4,5-טריפוספט (IP3). GPCRs המצומדים ל- Gα12/13 מפעילים את גורמי החלפת הנוקלאוטידים RhoGTPase 5,6. GPCRs הם המטרה של יותר מ -50% של תרופות אנושיות ו acaricide, amitraz4. כאשר GPCRs מתמרים אותות כה מגוונים, הם מהווים יעדים מבטיחים לפיתוח חומרי הדברה חדשניים המשבשים תפקודים פיזיולוגיים ספציפיים לחסרי חוליות.
המטרה של HTS היא לזהות מולקולות פגיעות שיכולות לווסת את תפקודי הקולטן. HTS כולל פיתוח בדיקות, מזעור ואוטומציה7. GPCRs נוירופפטידים של פרוקי רגליים מעורבים ברוב הפונקציות הפיזיולוגיות, כגון התפתחות, נפילה ואקדיזיס, הפרשה, גיוס אנרגיה ורבייה4. רוב הנוירופפטידים GPCRs של פרוקי רגליים ומטזואנים מאותתים דרך מפל איתות הסידן 2,6,8,9,10, כגון בקולטני הפפטיד המיואינהימבורי וה-SIFamide של הקרצייה השחורה Ixodes scapularis; הליגנדות שלהם הן אנטגוניסטיות במבחני תנועתיות אחוריים, כאשר SIF מעורר התכווצות ו- MIP מעכב אותו11,12. קולטן דמוי NPY של יתוש הקדחת הצהובה, Aedes aegypti, מווסת את נקבת הפונדקאית המחפשת13. בהשוואה למבחנים חלופיים אחרים לגיוס סידן, כגון בדיקת הסידן הביולומינסנציהaequorin 14, בדיקת הסידן הפלואורסצנטי קלה לביצוע, אינה דורשת העברה של חלבונים אחרים המזהים סידן רקומביננטי, והיא חסכונית. בדיקת הסידן הפלואורסצנטי מייצרת אות ממושך בהשוואה לאות הקינטי המהיר המתקבל בבדיקת הסידן הביולומינסנציה של אקוורין14,15.
בדוגמה כאן, קולטן קינין מקרציית קדחת הבקר, Rhipicephalus microplus, התבטא באופן רקומביננטי בקו התאים CHO-K1 ושימש לבדיקת סידן פלואורסצנטי. יש רק גן אחד של קולטן קינין שנמצא במיקרופלוס R. ; הקולטן מאותת דרך מסלול איתות תלוי חלבון Gq ומפעיל את השטף של Ca2+ ממאגרי סידן לתוך החלל התוך-תאי16. ניתן לזהות ולכמת את התהליך הזה על-ידי פלואורופור, שמעורר אות פלואורסצנטי בעת קשירת יוני סידן (איור 1).
קולטן קינין הוא GPCR ספציפי לחסרי חוליות, השייך לקולטנים דמויי רודופסין מסוג A. קינין הוא נוירופפטיד איתות עתיק שנמצא במולוסקה, סרטנאים, חרקים ואקרי 4,17,18. קולופטרנים (חיפושיות) חסרים את מערכת האיתות של קינין; אצל היתוש Aedes aegypti, יש רק קולטן קינין אחד שקושר שלושה אדסקינינים, בעוד שלדרוזופילה מלנוגסטר יש קולטן קינין אחד עם דרוסוקינין כליגנדייחודי 19,20,21. אין קינינים הומולוגיים או קולטני קינין אצל בעלי חוליות. למרות שהתפקוד המדויק של קינין אינו ידוע בקרציות, הנקבות המושתקות ב-RNAi של קולטן קינין של R. microplus מראות ירידה משמעותית בכושר הרבייה22. קינינים הם פפטידים פלוטרופיים בחרקים. ב Drosophila melanogaster, הם מעורבים הן במערכת ויסות העצבים המרכזית והן ההיקפית 23, טרום אקדיזה24, האכלה 25, מטבוליזם 26, ודפוסי פעילות שינה26,27, כמו גם תנועה זחל 28. קינינים מווסתים את ההתכווצות האחורית, דיאורזיס והזנה ביתוש A. aegypti 29,30,31. לפפטידים של קינין יש פנטפפטיד C-terminal Phe-X1-X2-Trp-Gly-NH2 שמור, שהוא הרצף המינימלי הנדרש לפעילות ביולוגית32. הספציפיות של פרוקי הרגליים, הגודל הקטן של הליגנד האנדוגני, מה שהופך אותם לנוחים להפרעות של מולקולות קטנות, והתפקודים הפליאוטרופיים בחרקים הופכים את קולטן קינין למטרה מבטיחה להדברה4.
מבחן “התוספת הכפולה” (איור 2) מאפשר זיהוי של אגוניסטים או אנטגוניסטים באותה בדיקת HTS15. הוא מותאם מבדיקת “תוספת כפולה” המשמשת בדרך כלל בתעשיית התרופות לגילוי תרופות33. בקצרה, התוספת הראשונה של תרופות לצלחת התא מאפשרת זיהוי של אגוניסטים פוטנציאליים בספרייה הכימית כאשר אות פלואורסצנטי גבוה יותר מזוהה בהשוואה ליישום של בקרת הממס. לאחר 5 דקות של דגירה עם מולקולות קטנות אלה, אגוניסט ידוע (פפטיד קינין) מוחל על כל הבארות. אותן בארות שקיבלו באופן אקראי אנטגוניסט מצלחת התרופה מציגות אות פלואורסצנטי נמוך יותר על תוספת אגוניסטית בהשוואה לבארות הבקרה שקיבלו את הממס בתוספת הראשונה. בדיקה זו מאפשרת זיהוי של אגוניסטים ואנטגוניסטים פוטנציאליים עם אותם תאים. בפרויקט HTS סטנדרטי, מולקולות פגיעות אלה יאומתו עוד יותר באמצעות מבחני מינון-תגובה ועל ידי מבחני פעילות ביולוגית נוספים, שאינם מוצגים כאן.
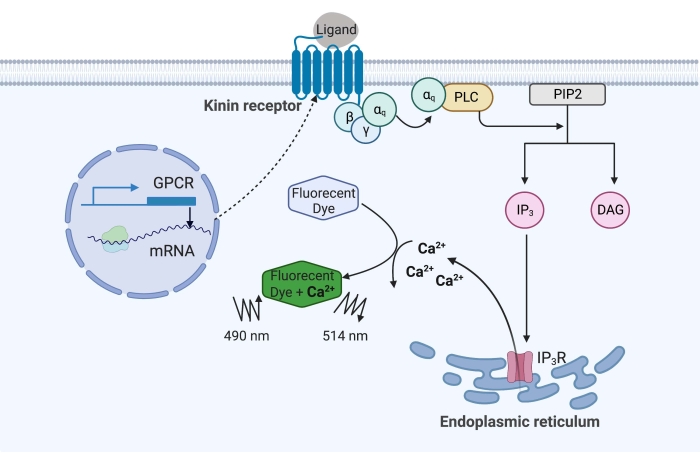
איור 1: המחשה של מנגנון בדיקת הסידן הפלואורסצנטי. חלבון ה-Gq מפעיל את מסלול איתות הסידן התוך-תאי. קולטן קינין (קולטן מצומד לחלבון G) בא לידי ביטוי רקומביננטי בתאי CHO-K1. כאשר הליגנד האגוניסט נקשר לקולטן, חלבון ה-Gq הקשור לקולטן קינין מפעיל את PLC, מה שמזרז את ההמרה של מולקולת PIP2 ל-IP3 ו-DAG. לאחר מכן, IP 3 נקשר ל-IP3R על פני השטח של הרשתית האנדופלסמית, מה שמוביל לשחרור Ca 2+ לתוך הציטופלסמה, שם יוני Ca2+ נקשרים לפלואורופורים ומעוררים אות פלואורסצנטי. ניתן לקבל את אות הפלואורסצנציה על ידי עירור ב 490 ננומטר ולזהות ב 514 ננומטר. קיצורים: GPCR = קולטן מצומד חלבון G; PLC = פוספוליפאז C; PIP2 = פוספטידילינוזיטול 4,5-ביספוספט; IP3 = אינוזיטול טריספוספט; DAG = דיאצילגליצרול; IP3 R = קולטן IP3. נוצר באמצעות BioRender.com. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
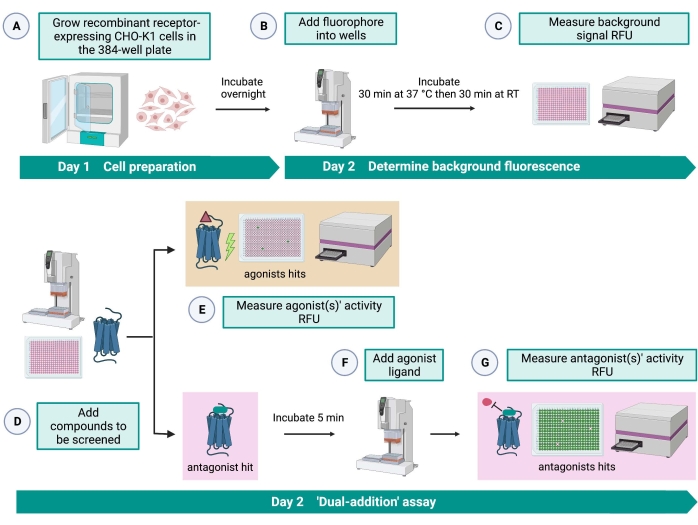
איור 2: תהליך העבודה של הקרנה בתפוקה גבוהה של מולקולות קטנות על קולטן מצומד לחלבון G המתבטא בתאי CHO-K1. (A) תאי CHO-K1 רקומביננטיים המבטאים ביציבות את קולטן הקינין נוספו לצלחת 384 באר (10,000 תאים/באר) באמצעות מערכת טיפול בנוזל (25 μL/well) ודוגרו באינקובטור CO2 לח במשך 12-16 שעות. (ב ) חיץ הבדיקה המכיל את הצבע הפלואורסצנטי (25 μL/well) נוסף ללוחית התא באמצעות מערכת טיפול בנוזל. הצלחת דוגרה במשך 30 דקות בטמפרטורה של 37 מעלות צלזיוס למשך 30 דקות ושיוותה ב-RT למשך 30 דקות נוספות. (D) תמיסות סמים מצלחת ספרייה של 384 בארות וממס ריק (כולם ב-0.5 μL/well) נוספו ללוחית הבדיקה התאית באמצעות מערכת טיפול בנוזלים. (E) תגובות פלואורסצנטיות של סידן תאי נמדדו עם קורא הצלחות מיד לאחר הוספת תמיסות התרופה; תרכובות המעוררות אותות פלואורסצנטיים גבוהים מהממוצע נבחרו כפגיעות אגוניסטיות. להיטים אנטגוניסטיים שחוסמים את ה-GPCR (אייקון למטה) נחשפו לאחר הוספת האגוניסט הפפטידי במהלך שלב G. (F) באותה צלחת בדיקה, לאחר 5 דקות של דגירה של התאים עם תרכובות הקרנה, פפטיד אגוניסט אנדוגני Rhimi-K-1 (QFSPWGamide) של קולטן קינין קרציות נוסף לכל באר (1 μM). (G) תגובות פלואורסצנטיות תאיות לאחר תוספת הפפטיד האגוניסטית נמדדו על ידי קורא הלוחות באופן מיידי. תרכובות המעכבות את האות הפלואורסצנטי נבחרו כפגיעות אנטגוניסטיות. קיצורים: GPCR = קולטן מצומד חלבון G; RT = טמפרטורת החדר; RFU = יחידות פלואורסצנטיות יחסיות. נוצר באמצעות BioRender.com. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
