Los receptores acoplados a proteínas G (GPCR), que están presentes desde la levadura hasta los humanos, representan la mayor superfamilia de receptores en muchos organismos1. Desempeñan un papel crítico en la regulación de casi todos los procesos biológicos en los animales. Hay 50-200 GPCR en el genoma de los artrópodos, lo que significa que representan la superfamilia de receptores de membrana más grande2. Se clasifican en seis clases principales, A-F, en función de su similitud de secuencia y funciones3. Los GPCR transducen diversas señales extracelulares, como las de hormonas, neuropéptidos, aminas biogénicas, glutamato, protones, lipoglicoproteínas y fotones4. Los GPCR se acoplan a las proteínas G heterotrímero (Gα, Gβ y Gγ) para transmitir señales aguas abajo. Los GPCR acoplados a las proteínas Gαs o Gαi/o aumentan o disminuyen, respectivamente, los niveles intracelulares de 3′, 5′ -monofosfato de adenosina cíclico (cAMP) mediante la activación o inhibición de la adenilil ciclasa. Los GPCR acoplados a Gαq/11 inducen la liberación de calcio de las reservas de calcio del retículo endoplásmico mediante la activación de la vía de la fosfolipasa C (PLC)-inositol-1,4,5-trifosfato (IP3). Los GPCR acoplados a Gα12/13 activan los factores de intercambio de nucleótidos RhoGTPasa 5,6. Los GPCR son el objetivo de más del 50% de los medicamentos humanos y un acaricida, amitraz4. A medida que los GPCR transducen señales tan diversas, son objetivos prometedores para desarrollar nuevos pesticidas que interrumpen las funciones fisiológicas específicas de los invertebrados.
El objetivo de HTS es identificar moléculas de impacto que puedan modular las funciones del receptor. HTS implica el desarrollo de ensayos, miniaturización y automatización7. Los GPCR del neuropéptido artrópodo están involucrados en la mayoría de las funciones fisiológicas, como el desarrollo, la muda y la ecdisis, la excreción, la movilización de energía y la reproducción4. La mayoría de los GPCR neuropeptídicos de artrópodos y metazoos señalan a través de la cascada de señalización de calcio 2,6,8,9,10, como en el péptido mioinhibitorio y los receptores SIFamida de la garrapata de patas negras Ixodes scapularis; sus ligandos son antagónicos en los ensayos de motilidad del intestino posterior, con SIF provocando contracción y MIP inhibiéndola11,12. Un receptor similar al NPY del mosquito de la fiebre amarilla, Aedes aegypti, regula la búsqueda de huéspedes femeninos13. En comparación con otros ensayos alternativos de movilización de calcio, como el ensayo de bioluminiscencia cálcica aequorina14, el ensayo de fluorescencia de calcio es fácil de realizar, no requiere la transfección de otras proteínas detectoras de calcio recombinantes y es rentable. El ensayo de fluorescencia de calcio produce una señal prolongada en comparación con la señal cinética rápida obtenida en el ensayo de bioluminiscencia cálcica aequorina14,15.
En el ejemplo aquí, el receptor de cinina de la garrapata de la fiebre del ganado, Rhipicephalus microplus, se expresó recombinantemente en la línea celular CHO-K1 y se utilizó para el ensayo de fluorescencia de calcio. Solo hay un gen receptor de cinina que se encuentra en R. microplus; el receptor señala a través de una vía de señalización dependiente de la proteína Gq y desencadena el flujo de Ca2+ de las reservas de calcio al espacio intracelular16. Este proceso puede ser detectado y cuantificado por un fluoróforo, que provoca una señal de fluorescencia al unir iones de calcio (Figura 1).
El receptor de quinina es un GPCR específico de invertebrados, que pertenece a los receptores similares a la rodopsina de clase A. La quinina es un antiguo neuropéptido de señalización que está presente en moluscos, crustáceos, insectas y acari 4,17,18. Los coleópteros (escarabajos) carecen del sistema de señalización de cininas; en el mosquito Aedes aegypti, solo hay un receptor de quinina que se une a tres aedeskininas, mientras que Drosophila melanogaster tiene un receptor de cinina con drosokinina como ligando único 19,20,21. No hay cininas homólogas o receptores de cinina en vertebrados. Aunque la función exacta de la cinina es desconocida en las garrapatas, las hembras silenciadas por ARNi del receptor de cinina de R. microplus muestran una aptitud reproductiva significativamente reducida22. Las cininas son péptidos pleotrópicos en insectos. En Drosophila melanogaster, están involucrados en los sistemas reguladores nerviosos central y periférico 23, pre-ecdisis24, alimentación 25, metabolismo 26 y patrones de actividad del sueño26,27, así como locomoción larvaria 28. Las cininas regulan la contracción del intestino posterior, la diuresis y la alimentación en el mosquito A. aegypti 29,30,31. Los péptidos de cinina tienen un pentapéptido C-terminal conservado Phe-X1-X2-Trp-Gly-NH2, que es la secuencia mínima requerida para la actividad biológica32. La especificidad de los artrópodos, el pequeño tamaño del ligando endógeno, que los hace susceptibles a la interferencia de moléculas pequeñas, y las funciones pleiotrópicas en los insectos hacen del receptor de cinina un objetivo prometedor para el control de plagas4.
El ensayo de “doble adición” (Figura 2) permite la identificación de agonistas o antagonistas en el mismo ensayo HTS15. Está adaptado de un ensayo de “doble adición” que se utiliza comúnmente en la industria farmacéutica para el descubrimiento de fármacos33. En resumen, la primera adición de fármacos en la placa celular permite la identificación de posibles agonistas en la biblioteca química cuando se detecta una señal de fluorescencia más alta en comparación con la aplicación del control de disolvente. Después de 5 minutos de incubación con estas moléculas pequeñas, se aplica un agonista conocido (péptido de cinina) a todos los pocillos. Aquellos pocillos que recibieron al azar un antagonista de la placa de fármaco muestran una señal de fluorescencia más baja tras la adición de agonistas en comparación con los pocillos de control que recibieron el solvente en la primera adición. Este ensayo permite la identificación de posibles agonistas y antagonistas con las mismas células. En un proyecto estándar de HTS, estas moléculas de éxito se validarían aún más a través de ensayos de dosis-respuesta y mediante ensayos de actividad biológica adicionales, que no se muestran aquí.
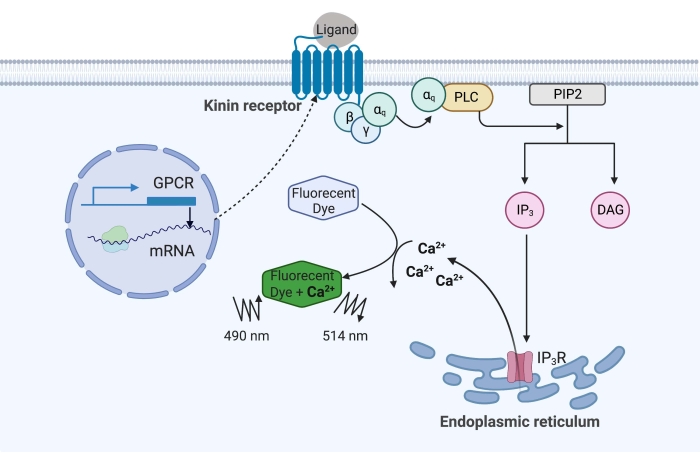
Figura 1: Ilustración del mecanismo de ensayo de fluorescencia de calcio. La proteína Gq desencadena la vía de señalización de calcio intracelular. El receptor de cinina (receptor acoplado a proteína G) se expresó recombinantemente en células CHO-K1. Cuando el ligando agonista se une al receptor, la proteína Gq asociada con el receptor de quinina activa PLC, que cataliza la conversión de una molécula PIP2 en IP3 y DAG. IP 3 luego se une al IP3R en la superficie del retículo endoplásmico, lo que lleva a la liberación de Ca 2+ en el citoplasma, donde los iones de Ca2+ se unen a los fluoróforos y provocan una señal de fluorescencia. La señal de fluorescencia puede obtenerse por excitación a 490 nm y detectarse a 514 nm. Abreviaturas: GPCR = receptor acoplado a proteína G; PLC = fosfolipasa C; PIP2 = fosfatidilinositol 4,5-bisfosfato; IP3 = trifosfato de inositol; DAG = diacilglicerol; IP3 R = receptor IP3. Creado con BioRender.com. Haga clic aquí para ver una versión más grande de esta figura.
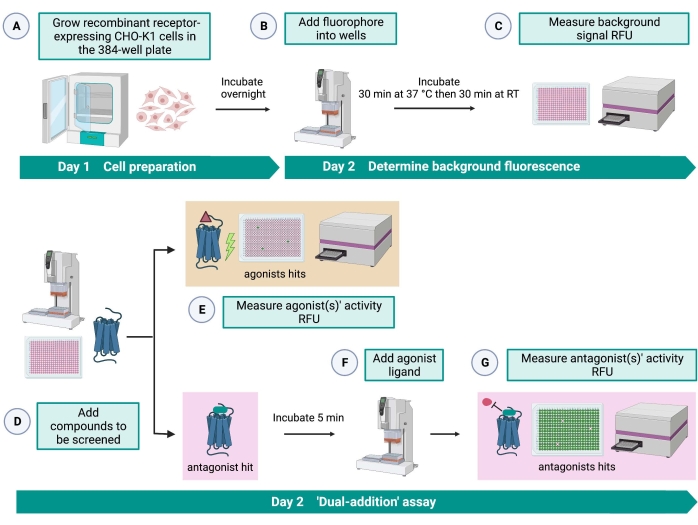
Figura 2: El flujo de trabajo para el cribado de alto rendimiento de moléculas pequeñas en un receptor acoplado a proteína G expresado en células CHO-K1. (A) Las células CHO-K1 recombinantes que expresan de manera estable el receptor de cinina se agregaron a la placa de 384 pocillos (10,000 células / pocillo) utilizando un sistema de manejo de líquidos (25 μL / pocillo) y se incubaron en una incubadora humidificada de CO2 durante12-16 h. (B ) El tampón de ensayo que contenía el colorante fluorescente (25 μL/pocillo) se añadió a la placa celular utilizando un sistema de manipulación de líquidos. La placa se incubó durante 30 min a 37 °C durante 30 min y se equilibró a RT durante otros 30 min. (C) La señal de fluorescencia de fondo de las células en cada pocillo se midió con un lector de placas. (D) Se agregaron soluciones farmacológicas de una placa de biblioteca de 384 pocillos y disolvente en blanco (todo a 0,5 μL/pocillo) a la placa de ensayo celular utilizando un sistema de manejo de líquidos. (E) Las respuestas de fluorescencia de calcio celular se midieron con el lector de placas inmediatamente después de la adición de las soluciones farmacológicas; Los compuestos que provocan señales de fluorescencia superiores a la media se seleccionaron como agonistas acertados. Los golpes antagonistas que bloquean el GPCR (icono a continuación) se revelaron después de la adición del agonista peptídico durante el paso G. (F) En la misma placa de ensayo, después de 5 minutos de incubación de las células con compuestos de cribado, se añadió un péptido agonista endógeno Rhimi-K-1 (QFSPWGamida) del receptor de quinina de garrapata a cada pocillo (1 μM). (G) Las respuestas de fluorescencia celular después de la adición del péptido agonista fueron medidas por el lector de placas inmediatamente. Los compuestos que inhiben la señal de fluorescencia se seleccionaron como antagonistas de los hits. Abreviaturas: GPCR = receptor acoplado a proteína G; RT = temperatura ambiente; RFU = unidades de fluorescencia relativa. Creado con BioRender.com. Haga clic aquí para ver una versión más grande de esta figura.
