Рецепторы, связанные с G-белком (GPCR), которые присутствуют от дрожжей к человеку, представляют собой крупнейшее надсемейство рецепторов во многих организмах1. Они играют решающую роль в регулировании почти всех биологических процессов у животных. В геноме членистоногих насчитывается 50-200 GPCR, что означает, что они представляют собой самое большое надсемейство мембранных рецепторов2. Они классифицируются на шесть основных классов, A-F, на основе их сходства последовательностей и функций3. GPCR трансдуцируют различные внеклеточные сигналы, такие как гормоны, нейропептиды, биогенные амины, глутамат, протон, липогликопротеины и фотоны4. GPCR соединяются с гетеротримерными G-белками (Gα, Gβ и Gγ) для передачи последующих сигналов. GPCR в сочетании с белками Gαs или Gαi/o увеличивают или уменьшают, соответственно, внутриклеточные 3′, 5′-циклические уровни аденозинмонофосфата (цАМФ) путем активации или ингибирования аденилциклазы. GPCR, связанные с Gαq/11, индуцируют высвобождение кальция из эндоплазматических ретикулумных запасов кальция путем активации пути фосфолипазы C (PLC)-инозитол-1,4,5-трифосфата (IP3). GPCR в сочетании с Gα12/13 активируют RhoGTPase нуклеотидные обменные факторы 5,6. GPCR являются мишенью более 50% лекарств для человека и акарицида, амитраза4. Поскольку GPCR передают такие разнообразные сигналы, они являются многообещающими целями для разработки новых пестицидов, которые нарушают физиологические функции, специфичные для беспозвоночных.
Целью HTS является идентификация пораженных молекул, которые могут модулировать функции рецепторов. HTS включает в себя разработку анализов, миниатюризацию и автоматизацию7. Членистоногие нейропептидные GPCR участвуют в большинстве физиологических функций, таких как развитие, линька и экдиз, экскреция, мобилизация энергии и размножение4. Большинство нейропептидных GPCR членистоногих и метазоев сигнализируют через кальциевый сигнальный каскад 2,6,8,9,10, например, в миоингибиторных пептидных и SIFамидных рецепторах черноногих клещей Ixodes scapularis; их лиганды антагонистичны в анализах моторики задней кишки, при этом SIF вызывает сокращение, а MIP ингибирует его11,12. NPY-подобный рецептор комара желтой лихорадки, Aedes aegypti, регулирует женщину-хозяина, ищущего13. По сравнению с другими альтернативными анализами мобилизации кальция, такими как анализ биолюминесценции кальция экворина14, анализ флуоресценции кальция прост в выполнении, не требует трансфекции других рекомбинантных белков, обнаруживающих кальций, и является экономически эффективным. Анализ флуоресценции кальция дает пролонгированный сигнал по сравнению с быстрым кинетическим сигналом, полученным в анализе биолюминесценции кальция экворина14,15.
В приведенном здесь примере кининовый рецептор от клеща лихорадки крупного рогатого скота, Rhipicephalus microplus, был рекомбинантно экспрессирован в клеточной линии CHO-K1 и использовался для анализа флуоресценции кальция. Существует только один ген рецептора кинина, обнаруженный в R. microplus; рецептор сигнализирует через Gq белок-зависимый сигнальный путь и запускает отток Ca2+ из запасов кальция во внутриклеточное пространство16. Этот процесс может быть обнаружен и количественно определен флуорофором, который вызывает флуоресцентный сигнал при связывании ионов кальция (рисунок 1).
Кининовый рецептор представляет собой специфический для беспозвоночных GPCR, который относится к родопсиноподобным рецепторам класса А. Кинин является древним сигнальным нейропептидом, который присутствует в моллюсках, ракообразных, насекомых и акари 4,17,18. У колеоптерянов (жуков) отсутствует кининовая сигнальная система; у комара Aedes aegypti есть только один рецептор кинина, который связывает три эдескинина, в то время как Drosophila melanogaster имеет один рецептор кинина с дросокинином в качестве уникального лиганда 19,20,21. У позвоночных нет гомологичных кининов или кининовых рецепторов. Хотя точная функция кинина у клещей неизвестна, у женщин R. microplus, заглушенных рецептором RNAi, значительно снижается репродуктивная пригодность22. Кинины являются плеотропными пептидами у насекомых. У Drosophila melanogaster они участвуют как в центральной, так и в периферической нервной регуляторной системах23, преэкдисисе24, питании25,метаболизме 26 и паттернах активности сна 26,27, а также личиночной локомоции28. Кинины регулируют сокращение задней кишки, диурез и питание у комара A. aegypti 29,30,31. Кининовые пептиды имеют законсервированный С-концевой пентапептид Phe-X1-X2-Trp-Gly-NH2, который является минимально необходимой последовательностью для биологической активности32. Специфичность членистоногих, малый размер эндогенного лиганда, который делает их поддающимися низкомолекулярной интерференции, и плейотропные функции у насекомых делают кининовый рецептор перспективной мишенью для борьбы с вредителями4.
Анализ «двойного сложения» (рисунок 2) позволяет идентифицировать агонисты или антагонисты в том же анализе HTS15. Он адаптирован из анализа «двойного добавления», который обычно используется в фармацевтической промышленности для открытия лекарств33. Короче говоря, первое добавление лекарств в клеточную пластину позволяет идентифицировать потенциальные агонисты в химической библиотеке при обнаружении более высокого флуоресцентного сигнала по сравнению с применением контроля растворителя. После 5 мин инкубации с этими небольшими молекулами на все скважины наносится известный агонист (кининовый пептид). Те скважины, которые случайным образом получили антагонист из лекарственной пластины, показывают более низкий флуоресцентный сигнал при добавлении агониста по сравнению с контрольными лунками, которые получили растворитель при первом добавлении. Этот анализ позволяет идентифицировать потенциальные агонисты и антагонисты с одними и теми же клетками. В стандартном проекте HTS эти пораженные молекулы будут дополнительно проверены с помощью анализов «доза-реакция» и дополнительных анализов биологической активности, которые здесь не показаны.
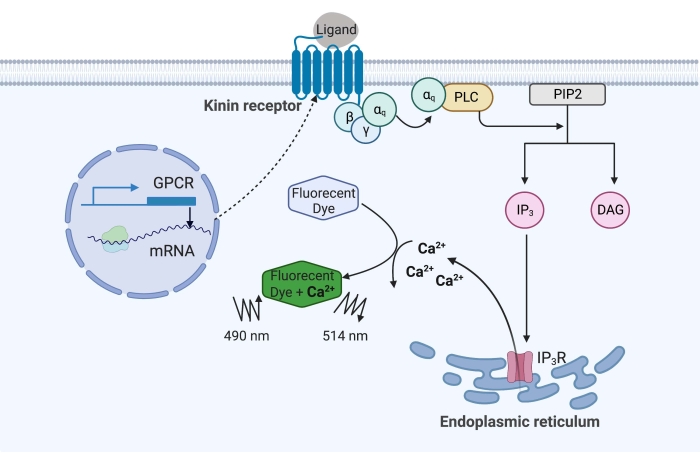
Рисунок 1: Иллюстрация механизма анализа флуоресценции кальция. Белок Gq запускает внутриклеточный сигнальный путь кальция. Рецептор кинина (рецептор, связанный с G-белком) был рекомбинантно экспрессирован в клетках CHO-K1. Когда лиганд агониста связывается с рецептором, белок Gq, связанный с рецептором кинина, активирует PLC, который катализирует превращение молекулы PIP2 в IP3 и DAG. Затем IP3 связывается с IP3R на поверхности эндоплазматического ретикулума, что приводит к высвобождению Ca2+ в цитоплазму, где ионы Ca2+ связываются с флуорофорами и вызывают флуоресцентный сигнал. Флуоресцентный сигнал может быть получен возбуждением при 490 нм и обнаружен при 514 нм. Сокращения: GPCR = рецептор, связанный с G-белком; PLC = фосфолипаза C; PIP2 = фосфатидилинозитол 4,5-бисфосфат; IP3 = инозитолтрисфосфат; DAG = диацилглицерин; IP3R = IP3 рецептор. Создано с помощью BioRender.com. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
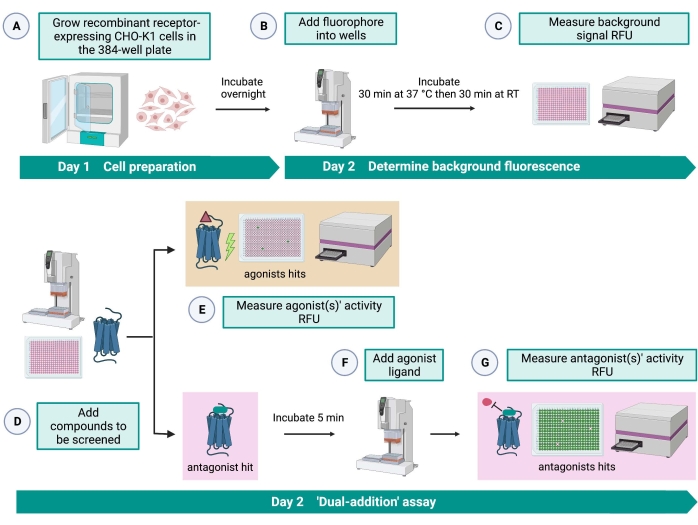
Рисунок 2: Рабочий процесс для высокопроизводительного скрининга малых молекул на рецепторе, связанном с G-белком, экспрессируемом в клетках CHO-K1. (A) Рекомбинантные клетки CHO-K1, стабильно экспрессирующие рецептор кинина, добавляли к 384-луночной пластине (10 000 клеток/лунка) с помощью жидкостной системы обработки (25 мкл/лунка) и инкубировали в увлажненном инкубатореCO2 в течение 12-16 ч. (B) ) Буфер анализа, содержащий флуоресцентный краситель (25 мкл/лунка), добавляли в клеточную пластину с помощью системы обработки жидкости. Пластину инкубировали в течение 30 мин при 37 °C в течение 30 мин и уравновешивали при RT еще 30 мин. (C) Фоновый флуоресцентный сигнал клеток в каждой скважине измеряли с помощью пластинчатого считывателя. (D) Растворы лекарственных средств из 384-луночной библиотечной пластины и пустого растворителя (все при 0,5 мкл/лунку) добавляли в пластину клеточного анализа с использованием системы обработки жидкости. (E) Клеточные реакции флуоресценции кальция измеряли с помощью считывателя пластин сразу после добавления растворов лекарственного средства; соединение (соединения), вызывающее сигналы флуоресценции выше среднего, были выбраны в качестве агониста удара (ударов). Удары антагониста, блокирующие GPCR (значок ниже), были выявлены после добавления агониста пептидов во время шага G. (F) В той же пробирной пластине после 5 мин инкубации клеток с экранирующими соединениями к каждой лунке добавляли эндогенный агонистический пептид Rhimi-K-1 (QFSPWGamide) клещевого кининового рецептора (1 мкМ). (G) Клеточные флуоресцентные реакции после добавления агониста пептида немедленно измерялись считывателем пластин. Соединение(я), ингибирующее флуоресцентный сигнал, было выбрано в качестве удара(ов) антагониста. Сокращения: GPCR = рецептор, связанный с G-белком; RT = комнатная температура; РФС = относительные единицы флуоресценции. Создано с помощью BioRender.com. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
