G-proteinkoblede receptorer (GPCR’er), som er til stede fra gær til mennesker, repræsenterer den største superfamilie af receptorer i mange organismer1. De spiller afgørende roller i reguleringen af næsten alle biologiske processer hos dyr. Der er 50-200 GPCR’er i leddyrs genom, hvilket betyder, at de repræsenterer den største membranreceptor-superfamilie2. De er klassificeret i seks hovedklasser, A-F, baseret på deres sekvenslighed og funktioner3. GPCR’er transducerer forskellige ekstracellulære signaler, såsom hormoner, neuropeptider, biogene aminer, glutamat, proton, lipoglycoproteiner og fotoner4. GPCR’er kobles til heterotrimer G-proteiner (Gα, Gβ og Gγ) for at transmittere nedstrømssignaler. GPCR’er koblet til Gαs eller Gαi / o-proteiner øger eller reducerer henholdsvis de intracellulære 3′, 5′-cykliske adenosinmonophosphatniveauer (cAMP) ved at aktivere eller hæmme adenylylcyclase. GPCR’er koblet til Gαq/11 inducerer calciumfrigivelse fra de endoplasmatiske retikulumcalciumlagre ved at aktivere phospholipase C (PLC)-inositol-1,4,5-triphosphat (IP3) vejen. GPCR’er koblet til Gα12/13 aktiverer RhoGTPase-nukleotidbyttefaktorer 5,6. GPCR’er er målet for mere end 50% af humane lægemidler og et acaricid, amitraz4. Da GPCR’er transducerer så forskellige signaler, er de lovende mål for udvikling af nye pesticider, der forstyrrer hvirvelløse dyr-specifikke fysiologiske funktioner.
Målet med HTS er at identificere hitmolekyler, der kan modulere receptorfunktioner. HTS involverer analyseudvikling, miniaturisering og automatisering7. Arthropod neuropeptid GPCR’er er involveret i de fleste fysiologiske funktioner, såsom udvikling, smeltning og ecdysis, udskillelse, energimobilisering og reproduktion4. De fleste neuropeptid-GPCR’er af leddyr og metazoer signalerer gennem calciumsignalkaskaden 2,6,8,9,10, såsom i myohæmmende peptid- og SIFamidreceptorer i den sortbenede flåt Ixodes scapularis; deres ligander er antagonistiske i bagtarmmotilitetsanalyser, hvor SIF fremkalder sammentrækning og MIP hæmmer det11,12. En NPY-lignende receptor af gul feber myg, Aedes aegypti, regulerer kvindelig vært søger13. Sammenlignet med andre alternative calciummobiliseringsassays såsom aequorin-calciumbioluminescensanalysen14 er calciumfluorescensanalysen let at udføre, kræver ikke transfektion af andre rekombinante calciumdetekterende proteiner og er omkostningseffektiv. Calciumfluorescensanalysen frembringer et forlænget signal sammenlignet med det hurtige kinetiske signal opnået i aequorincalciumbioluminescensanalysen14,15.
I eksemplet her blev kininreceptoren fra kvægfeberflåten, Rhipicephalus microplus, rekombinant udtrykt i CHO-K1-cellelinjen og anvendt til calciumfluorescensassayet. Der findes kun ét kininreceptorgen i R. microplus; receptoren signalerer gennem en Gq-proteinafhængig signalvej og udløser udstrømningen af Ca2+ fra calciumlagre til det intracellulære rum16. Denne proces kan detekteres og kvantificeres af en fluorofor, som fremkalder et fluorescenssignal ved binding af calciumioner (figur 1).
Kininreceptoren er en hvirvelløse dyr-specifik GPCR, som tilhører klasse A Rhodopsin-lignende receptorer. Kinin er et gammelt signalneuropeptid, der er til stede i Mollusca, Crustacea, Insecta og Acari 4,17,18. Coleopterans (biller) mangler kinin signalsystemet; i myggen Aedes aegypti er der kun en kininreceptor, der binder tre aedeskininer, mens Drosophila melanogaster har en kininreceptor med drosokinin som en unik ligand 19,20,21. Der er ingen homologe kininer eller kininreceptorer hos hvirveldyr. Selvom kinins nøjagtige funktion er ukendt i flåter, viser kininreceptorens RNAi-tavse hunner af R. microplus signifikant reduceret reproduktionsegnethed22. Kininer er pleotropiske peptider i insekter. I Drosophila melanogaster er de involveret i både de centrale og perifere nervereguleringssystemer23, præ-ecdysis24, fodring25, stofskifte 26 og søvnaktivitetsmønstre 26,27 samt larvebevægelse 28. Kinins regulerer hindgut sammentrækning, diurese og fodring i myggen A. aegypti 29,30,31. Kininpeptiderne har et konserveret C-terminalt pentapeptid Phe-X1-X2-Trp-Gly-NH2, som er den mindste krævede sekvens for biologisk aktivitet32. Arthropodspecificiteten, den lille størrelse af den endogene ligand, som gør dem modtagelige for småmolekyleinterferens, og de pleiotropiske funktioner i insekter gør kininreceptoren til et lovende mål for skadedyrsbekæmpelse4.
“Dual-addition”-assayet (figur 2) gør det muligt at identificere agonister eller antagonister i samme HTS-assay15. Det er tilpasset fra et “dual-addition” assay, der almindeligvis anvendes i medicinalindustrien til lægemiddelopdagelse33. Kort sagt tillader den første tilsætning af lægemidler i cellepladen identifikation af potentielle agonister i det kemiske bibliotek, når der detekteres et højere fluorescenssignal sammenlignet med anvendelsen af opløsningsmiddelkontrollen. Efter 5 minutters inkubation med disse små molekyler påføres en kendt agonist (kininpeptid) på alle brøndene. De brønde, der tilfældigt modtog en antagonist fra lægemiddelpladen, viser et lavere fluorescenssignal ved agonisttilsætning sammenlignet med kontrolhullerne, der modtog opløsningsmidlet i den første tilsætning. Dette assay tillader derefter identifikation af potentielle agonister og antagonister med de samme celler. I et standard HTS-projekt ville disse hitmolekyler blive yderligere valideret gennem dosis-respons-assays og ved yderligere biologiske aktivitetsassays, som ikke er vist her.
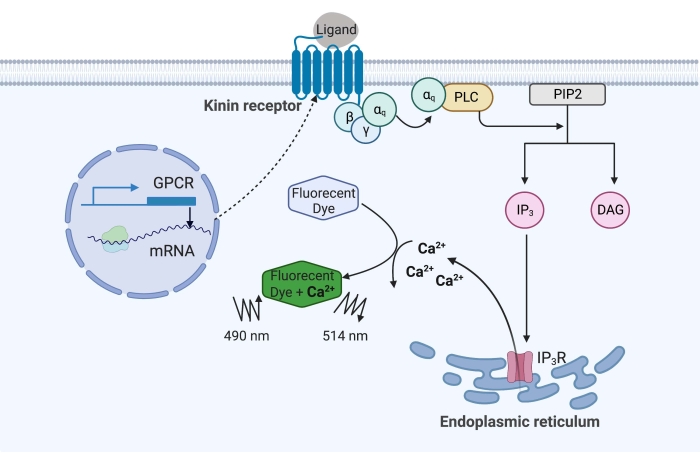
Figur 1: Illustration af calciumfluorescensanalysemekanismen. Gq-proteinet udløser den intracellulære calciumsignalvej. Kininreceptoren (G-proteinkoblet receptor) blev rekombinant udtrykt i CHO-K1-celler. Når agonistenaganden binder til receptoren, aktiverer Gq-proteinet, der er forbundet med kininreceptoren, PLC, som katalyserer omdannelsen af et PIP2-molekyle til IP3 og DAG. IP 3 binder derefter til IP3R på overfladen af det endoplasmatiske retikulum, hvilket fører til frigivelse af Ca 2+ i cytoplasmaet, hvor Ca2+ ioner binder til fluoroforerne og fremkalder et fluorescenssignal. Fluorescenssignalet kan opnås ved excitation ved 490 nm og detekteres ved 514 nm. Forkortelser: GPCR = G proteinkoblet receptor; PLC = phospholipase C; PIP2 = phosphatidylinositol-4,5-bisphosphat; IP3 = inositoltrisphosphat; DAG = diacylglycerol; IP3 R = IP3 receptor. Oprettet med BioRender.com. Klik her for at se en større version af denne figur.
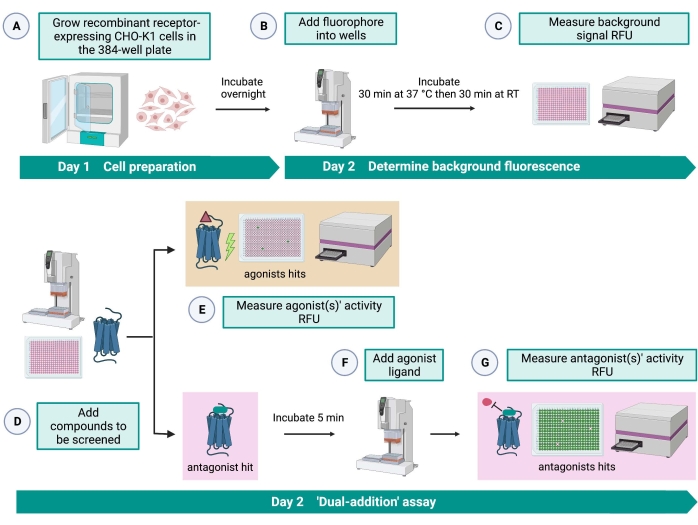
Figur 2: Arbejdsgangen for high-throughput screening af små molekyler på en G-proteinkoblet receptor udtrykt i CHO-K1-celler. (A) Rekombinante CHO-K1-celler, der stabilt udtrykker kininreceptoren, blev tilsat til 384-brøndpladen (10.000 celler / brønd) ved hjælp af et væskehåndteringssystem (25 μL / brønd) og inkuberet i en befugtet CO2 -inkubator i 12-16 timer. (B ) Analysebufferen indeholdende det fluorescerende farvestof (25 μL/hul) blev tilsat cellepladen ved hjælp af et væskehåndteringssystem. Pladen blev inkuberet i 30 minutter ved 37 °C i 30 minutter og ekvilibreret ved RT i yderligere 30 minutter. (C) Baggrundsfluorescenssignalet fra cellerne i hvert hul blev målt med en pladelæser. (D) Lægemiddelopløsninger fra en 384-brønds biblioteksplade og blindopløsningsmiddel (alle ved 0,5 μL / brønd) blev tilsat til den cellulære analyseplade ved hjælp af et væskehåndteringssystem. (E) Cellulære calciumfluorescensresponser blev målt med pladelæseren umiddelbart efter tilsætning af lægemiddelopløsningerne; Forbindelse(r), der fremkaldte fluorescenssignaler, der var højere end gennemsnittet, blev udvalgt som agonisthits. Antagonisthits, der blokerer GPCR (ikonet nedenfor) blev afsløret efter tilsætning af peptidagonisten under trin G. (F) I samme analyseplade blev der efter 5 minutters inkubation af cellerne med screeningsforbindelser tilsat et endogent agonistpeptid Rhimi-K-1 (QFSPWGamid) af flåtkininreceptoren til hvert hul (1 μM). (G) Cellulære fluorescensresponser efter tilsætning af agonistpeptid blev straks målt af pladelæseren. Forbindelse(r), der hæmmer fluorescenssignalet, blev valgt som antagonisthits. Forkortelser: GPCR = G proteinkoblet receptor; RT = stuetemperatur; RFU = relative fluorescensenheder. Oprettet med BioRender.com. Klik her for at se en større version af denne figur.
