Tette barrierer skiller sentralnervesystemet (CNS) fra periferien, inkludert blod-hjernebarrieren (BBB) og blod-cerebrospinalvæsken (CSF) barrieren. Disse barrierene beskytter CNS mot eksterne fornærmelser og sikrer et balansert og kontrollert mikromiljø 1,2,3. Mens BBB har blitt grundig studert over tid, har blod-CSF-barrieren som ligger ved choroid plexus (CP) bare fått økende forskningsinteresse i løpet av det siste tiåret. Denne sistnevnte barrieren finnes i hjernens fire ventrikler (figur 1A, B) og er preget av et enkelt lag av choroid plexus epitelceller (CPE) som omgir en sentral stroma, lekkende kapillærer, fibroblaster og en lymfoid og myeloid cellepopulasjon (figur 1C) 4,5,6. CPE-cellene er godt forbundet med stramme kryss, og forhindrer dermed lekkasje fra de underliggende fenestrerte blodkapillærene inn i CSF og hjernen. I tillegg reguleres transport over CPE-cellene av en rekke transportsystemer innover og utover som styrer tilstrømningen av gunstige forbindelser (f.eks. næringsstoffer og hormoner) fra blodet til CSF og utstrømningen av skadelige molekyler (f.eks. metabolsk avfall, overflødige nevrotransmittere) i den andre retningen 1,6. For å kunne utøve sin aktive transportfunksjon inneholder CPE-cellene mange mitokondrier i deres cytoplasma7. Videre er CP den viktigste kilden til CSF og fungerer som gatekeeper av hjernen ved tilstedeværelse av bosatt inflammatoriske celler1. På grunn av sin unike beliggenhet mellom blodet og hjernen, er CP også perfekt posisjonert for å utføre immunovervåking8.
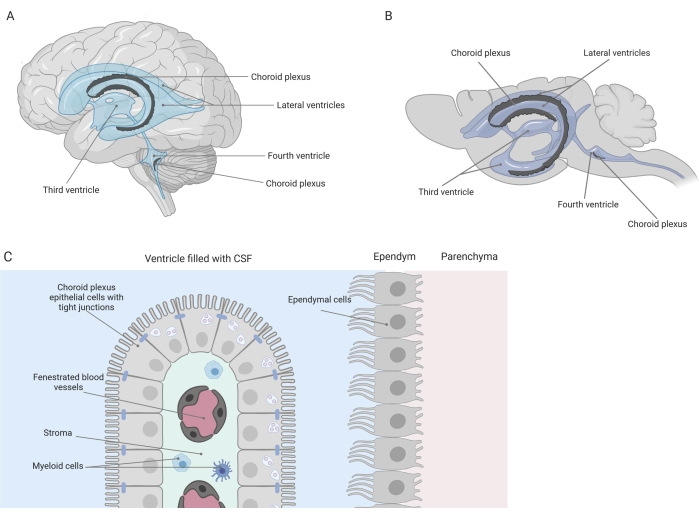
Figur 1 Skjematisk oversikt over lokalisasjon og sammensetning av choroid plexus (CP). (A,B) CP-vev finnes i de to laterale, tredje og fjerde ventriklene til (A) menneskelige og (B) musehjerner. (C) CP-vevet består av et enkelt lag av tett koblede kuboidale CP-epitelceller (CPE) som omgir fenestrerte kapillærer, løst bindevev og lymfoide og myeloide celler, og danner blod-cerebrospinalvæskebarrieren (tilpasset og modifisert fra referanse23). Figur opprettet med Biorender.com. Klikk her for å se en større versjon av denne figuren.
I løpet av det siste tiåret har økende bevis, inkludert flere rapporter fra vår forskningsgruppe, avslørt at CP spiller en sentral rolle i helse og sykdom 9,10,11,12,13,14,15,16,17,18 . For eksempel er det kjent at den aldrende blod-CSF-barrieren viser morfologiske endringer i blant annet kjernene, mikrovilli og kjellermembranen 1,19. I tillegg, i sammenheng med Alzheimers sykdom, er den generelle barriereintegriteten kompromittert, og alle disse aldersrelaterte endringene ser ut til å være enda mer uttalt 1,8,20. I tillegg til morfologiske endringer endres transkriptomet, proteomet og sekretomet til CP under sykdom 12,21,22,23. Dermed er avansert kunnskap om CP avgjørende for å bedre forstå sin rolle i nevrologiske sykdommer og potensielt utvikle nye terapeutiske strategier.
En effektiv metode for nøyaktig mikrodisseksjon av CP ut av hjerneventriklene er det første uvurderlige skrittet for å tillate riktig undersøkelse av denne lille hjernestrukturen. På grunn av sin sterkt vaskulariserte natur (figur 2B) kan CP som flyter inne i hjernens ventrikulære hulrom identifiseres ved hjelp av et kikkertmikroskop. Imidlertid er transkardial perfusjon ofte nødvendig for nedstrømsanalyse, noe som kompliserer riktig identifisering og isolering av CP-vevet (figur 2C). Hvis de videre behandlingstrinnene tillater det (f.eks. ved RNA- og proteinanalyse), kan CP visualiseres via transkardial perfusjon med bromfenolblått (figur 2A). Flere publikasjoner beskriver allerede isoleringen av CP fra rotte24 og musevalpehjerner25. Her beskrives en mikrodisseksjonsisolasjonsteknikk for å isolere CP fra voksne mus. Det er viktig at denne isolasjonsteknikken bevarer levedyktigheten, funksjonen og strukturen til cellene i CP. Isoleringen av CP flytende i fjerde og laterale ventrikler er beskrevet her. Kort sagt, musene er terminalt bedøvet og om nødvendig transkardialt perfusert. Det skal imidlertid bemerkes at perfusjon kan skade strukturen til cellene i CP. Følgelig, hvis prøven skal analyseres ved hjelp av transmisjonselektronmikroskopi (TEM), seriell blokk ansiktsskanning elektronmikroskopi (SBF-SEM), eller fokusert ionstråle SEM (FIB-SEM), bør perfusjon ikke utføres. Deretter isoleres hele hjernen, og tang brukes til å sagittalt hemisect hjernen. Herfra kan CP-ene som flyter i laterale ventrikler identifiseres og dissekeres, mens CP fra fjerde ventrikkel kan isoleres fra hjernesiden.
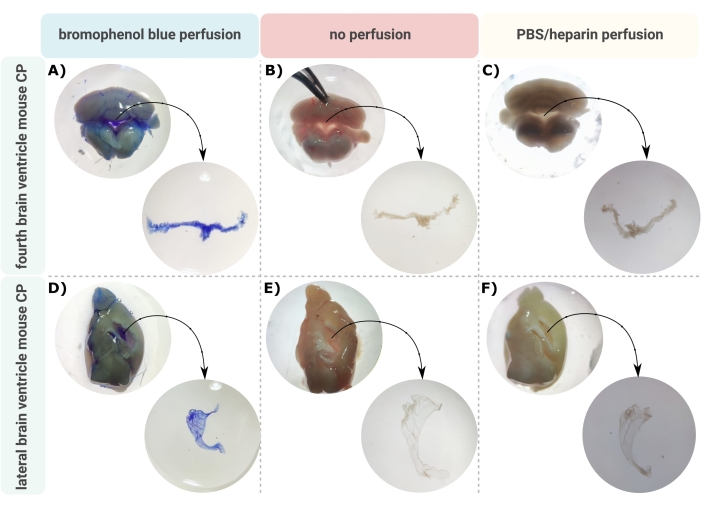
Figur 2 Visualisering av (A-C) fjerde og (D-F) laterale ventrikkel choroid plexus (CP) etter (A,D) bromfenolblå perfusjon, (B,E) ingen perfusjon og (C,F) perfusjon med PBS/heparin. Bildene er tatt med stereomikroskop (8x-32x forstørrelse). Klikk her for å se en større versjon av denne figuren.
Når CP er riktig dissekert ut av hjerneventriklene, kan et helt repertoar av teknikker brukes for å få ytterligere forståelse av funksjonen til denne strukturen. For eksempel kan flowcytometri eller enkeltcelle RNA-sekvensering utføres for å kvantifisere og fenotypisk analysere de infiltrerende inflammatoriske cellene under visse sykdomstilstander26,27. I tillegg til den cellulære sammensetningen kan den molekylære sammensetningen av CP analyseres for å vurdere tilstedeværelsen av cytokiner og kjemokiner via enzymbundet immunosorbentanalyse (ELISA), immunoblot, eller gjennom samtidig analyse av flere cytokiner ved bruk av cytokinperlematrisen28. Videre kan transkriptom-, vaskulær-, immuncellehistologi- og sekretomanalyser utføres på de mikrodissekerte CP-eksplantene29. Her brukes skanning elektronmikroskopi (SEM) på hele mount CP for å få en samlet oversikt over CP-strukturen. SEM bruker en stråle av fokuserte elektroner til å skanne over overflaten og skape et bilde av overflatens topografi og sammensetning. Siden bølgelengden til elektroner er mye mindre enn lyset, er oppløsningen av SEM i nanometerområdet og overlegen til et lysmikroskop. Følgelig kan morfologiske studier på subcellulært nivå utføres via SEM. Kort fortalt overføres den dissekerte CP umiddelbart til et glutaraldehydholdig fikseringsmiddel for en fiksering over natten, etterfulgt av osmikyring og uranylacetatfarging. Prøvene blir deretter behandlet med bly aspartat flekk, dehydrert, og til slutt innebygd for avbildning.
Dermed letter denne protokollen effektiv isolering av CP fra musens hjerneventrikler, som kan analyseres videre ved hjelp av en rekke nedstrøms teknikker for å undersøke dens struktur og funksjon.
