

Summary
Geri dönüşüm endozomları endosomal tübüler ağın bir parçasıdır. Burada, GFP-STX13'i organel işaretleyici olarak kullanarak geri dönüşüm endozomlarının dinamiklerini ölçmek için bir yöntem sunuyoruz.
Abstract
Geri dönüşüm endozomları (RE' ler), tüm hücre tiplerinde erken/sıralama endozomlarından üretilen tübüler-veziküler organellerdir. Bu organeller, melanositler tarafından üretilen lizozomla ilgili bir organel olan melanozomların biyogenezinde önemli bir rol oynar. REs, melanosit spesifik kargoyu oluşumları sırasında prematüre melanozomlara teslim eder. Hermansky-Pudlak sendromunun birkaç mutantında gözlenen REs neslindeki tıkanıklık, cilt, saç ve göz hipopigmentasyonu ile sonuçlanır. Bu nedenle, REs'in dinamiklerini (sayı ve uzunluğa bakın) incelemek, bu organellerin normal ve hastalık koşullarındaki işlevini anlamak için yararlıdır. Bu çalışmada, yerleşik bir SNARE STX13 kullanarak RE dinamiklerini ölçmeyi amaçlıyoruz.
Introduction
Melanin pigmentlerinin biyosentezi, geleneksel lizozomlarla birlikte var olan melanosit spesifik lizozomla ilişkili bir organel (LRO) olan melanozomlarda görülür. Endosit sistemi, iyonlaştırıcı radyasyona karşı cilt rengi ve fotoproteksiyon için gerekli olan melanozomların biyogenezinde önemli bir rol oynar1,2,3. Bu işlem sırasında melanin sentezleme enzimleri erken/tasnif endozomları üzerinde sıralanır ve daha sonra geri dönüşüm endozomları (REs)4,5,6,7,8,9,10 adı verilen tübüler veya veziküler endozomlar aracılığıyla erken melanozomlara taşınır. Bu organellerin hedeflenmesi ve füzyonu, tamamen fonksiyonel pigmentli melanozomların olgunlaşmasını düzenler7,11,12,13,14. Bu organellerin oluşumundaki kusurlar veya bu organellere giden yük tasnifleri, Hermansky-Pudlak sendromunda gözlenen okülokütan albinizme ve diğer klinik fenotiplere neden olur15,16.
Burada REs'i incelemek ve analiz etmek için basit bir mikroskopi tabanlı tekniği açıklıyoruz. Bu yöntemde, geri dönüşüm endozomları17 ve melanositlerdeki endozomlar ve melanozomlar arasındaki döngüler üzerinde bulunan bir transmembran proteini olan Qa-SNARE Syntaxin (STX)13'den yararlandık12,18. Ayrıca, N-terminal yapılandırılmamış düzenleyici etki alanının (synn veya STX13Δ129) silinmesi, SNARE'nin melanozomlara doğru ileriye doğru kaçakçılık yolunu ölçen melanozomlarda sıkışmasını sağlar12. Çalışmalarımızda bilinen bir geri dönüşüm endosomal marker Rab GTPase (Rab)11 kullandık14,19. GFP-STX13WT, GFP-STX13Δ129, mCherry-Rab11 ve TYRP1 proteinlerinin vahşi tip melanositlerde floresan görüntülemesi ve ardından göreceli lokalizasyonlarının ölçülmesi, melanozomlara hedeflemelerinin yanı sıra RE'lerin doğasını ve dinamiklerini sağlayacaktır. Bu nedenle, bu, MELANOSITLERDEKI REs dinamiklerini görselleştirmek ve ölçmek için kullanılabilecek basit bir tekniktir.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Protokol, melanositlerin tohumlenmesini ve ardından plazmidlerin transfeksiyonunu içerir. Diğer adımlar, RE'lerin uzunluğunu ve sayısını ölçmek için hücrelerin sabitlenmesi, immünostaining, görüntüleme ve analizini içerir. Protokolün ayrıntılı açıklaması aşağıda verilmiştir.
1. Önceden işlenmiş kapak örtülerinde fare melanositlerinin tohumlaştırılması
- Cam kapakları bir Petri kabına (yani 35 mm'lik bir kapta 4 - 5) bodrum membran matris ortamıyla (tam RPMI ortamında 1:20: RPMI + %10 ısı inaktive FBS + 1x Glutamin + 1x Antibiyotik karışımı) kapağa örtün ve doku kültürü davlumbazında 15 dakika kurutun. Kullanmadan önce kapakları 1x PBS ile bir kez yıkayın.
- Vahşi tip fare melanositlerini (C57BL/6J'den melan-Ink4a-Arf-1, a/a, Ink4a-Arf-/- fareler, 20'de tanımlanmış ve Welcome Trust Fonksiyonel Genomik Hücre Bankası'nda mevcuttur) tam RPMI ortamı ile desteklenmiş bir Petri kabında (yani 35 veya 60 mm'lik bir tabak) muhafaza edin.
- Hücreleri 1x PBS ile iki kez yıkayın ve trypnizing hücreleri için 0,5 veya 1 mL Trypsin-EDTA (%0,25) çözeltisi ekleyin. Petri kabından ayırma için hücreleri 2 - 5 dakika boyunca 37 °C'de kuluçkaya yatırın.
- 1 - 2 mL tam RPMI ortamı ekleyin, askıya alın ve ardından hücreleri bir santrifüj tüpüne aktarın.
- Hücre süspansiyonu 4 °C, 376 x g'da 5 dakika santrifüj edin ve ardından peleti 1x PBS'de yeniden saklayın.
- Santrifüjleme adımını tekrarlayın ve hücreleri 1 mL tam RPMI ortamında yeniden biriktirin.
- Bodrum membran orta kaplamalı kapaklardaki hücreleri% 50-60 izdiahta (4 - 5 kapaklı bir 35 mm hücre kültürü çanağı üzerinde yaklaşık 6 x 105 hücre) tohumlayın. Plakalı hücre süspansiyonu için her zaman tam RPMI ortamında 200 nM PMA (5 μL çalışma stoğu 40 μM phorbol 12-myristate 13-asetat ekleyin) ekleyin.
- Tohumlamadan sonra, plakayı 37 ° C'de 12- 24 saat boyunca kuluçkaya yatırın.
2. STX13 plazmidleri ile hücrelerin transfeksiyon
- Aşağıdaki reaktifleri kullanın: pEGFP-C1-STX13WT ve pEGFP-C1-STX13Δ129 (12 olarak açıklanmıştır). mCherry-Rab11, Graça Raposo, Institut Curie, Paris'ten (referans19'da açıklanmıştır) nazik bir hediyeydi. ATCC'den (TA99) anti-TYRP1 antikor.
- 12 - 24 saat tohumlamadan sonra, lipid bazlı bir transfeksiyon reaktifi kullanarak hücreleri plazmidlerle transfectect. 35 mm'lik bir çanak için, bir mikrosantrifüj tüpünde 250 μL OPTI-MEM ortamında transfeksiyon reaktifinin 5 μL'sini alın ve 250 μL OPTI-MEM'de her plazmidden yaklaşık 200 ng alın.
- DNA ve transfeksiyon reaktifi içeren tüpleri 5 dakika kuluçkaya yatırın. Tekrarlanan pipetleme olmadan karıştırın (toplam hacim yaklaşık 500 μL olacaktır). Oda sıcaklığında 30 dakika kuluçkaya yatır. RT'de 30 dakika boyunca her 10 dakikada bir tüpün elle dokunmasını gerçekleştirin.
- Kuluçka sırasında, hücreleri 1x PBS ile iki kez, opti-MEM ile bir kez yıkayın ve ardından hücrelere 1 mL OPTI-MEM ekleyin.
- 30 dakikalık inkübasyon sonrası, transfeksiyon reaktif-DNA karışımını yemeği kapatarak damla yönünde hücrelere ekleyin.
- Hücreleri 37 °C'de 6 saat kuluçkaya yatırın. OPTI-MEM ortamını transfeksiyon reaktifi ile epire edin ve 200 nM PMA ile desteklenmiş eksiksiz RPMI ortamı ekleyin.
- Hücreleri 37 °C'de 48 saat kuluçkaya yatırın.
3. Hücrelerin sabitlenmesi
NOT: Aşağıdaki işlem doku kültürü başlığı dışında gerçekleştirilir.
- 48 saat transfeksiyondan sonra, hücreleri 1x PBS ile iki kez yıkayın ve ardından hücreleri% 3 formaldehit (1x PBS'de taze hazırlanmış) ile 30 dakika sabitler.
- Sabitlemeden sonra, hücreleri 1x PBS ile iki kez yıkayın ve kapak kapaklarını bir sonraki kullanıma kadar 1x PBS'de saklayın. Alternatif olarak, hücreler cam slaytlara monte edilebilir (aşağıya bakın) veya 4 °C'de saklanabilir.
4. Hücrelerin immünostaining
- Nemli bir oda hazırlayın: parafin filmi kesilmiş parçasını alüminyum folyo ile kaplı bir Petri kabındaki nemli filtre kağıdına yerleştirin.
- 25 μL primer antikor çözeltisi hazırlayın (1x PBS'de %0,2 saponin, 1x PBS'de %0,1 BSA ve 1x PBS'de %0,02 Sodyum azit). 1:200'lük bir seyreltmede antikor ekleyin. Bu çözeltiyi nemli haznedeki bir parafin filmine damla olarak ekleyin.
- Kapak sapını forsepsle dikkatlice kaldırın, birincil antikor boyama çözeltisinin bu damlasına ters çevirin ve ardından nemli odanın kapağını kapatın. Oda sıcaklığında 30 dakika kuluçkaya yatır.
- Benzer şekilde, ikincil antikor çözeltisini 1:500 seyreltmede hazırlayın ve nemli haznedeki kapak kapağının yanındaki parafin filmine yerleştirin. Çekirdeği boyamak için çözeltiye DAPI (1:20.000 ila 1:30.000) ekleyin.
- Forseps kullanarak, kapak kapağını birincil antikor çözeltisinden dikkatlice alın ve üç kez 1x PBS'ye (cam bir behere) batırın.
- Kapak üzerindeki fazla PBS'yi çıkarmak için kağıt mendil üzerindeki kapak kılıfı'na dokunun. Nemli hazneye ikincil antikor boyama çözeltisine yerleştirin ve çözeltide floresan etiketli antikorların varlığı nedeniyle ışığa maruz bırakmayın.
- Kapak kapağını oda sıcaklığında 30 dakika boyunca tekrar kuluçkaya yatırın. Lütfen her zaman bu adımlar boyunca hücreleri olan kapak kılıfının yan tarafını not edin.
- Kuluçkadan sonra, kapak kapağını ikincil antikor çözeltisinden dikkatlice alın ve ardından 1x PBS'ye üç kez batırın. Ayrıca, kapak üzerindeki fazla PBS'yi çıkarmak için kağıt mendil üzerindeki kapak kılıfı'na dokunun.
- 12 μL Floromount-G montaj reaktifini bir cam kaydırağa yerleştirin ve lekeli kapak kapağını (cama doğru bakacak şekilde) montaj reaktifine dikkatlice yerleştirin. Cam slaydı kağıt mendile ters çevirin ve sonra hafifçe bastırın.
5. Hücrelerin floresan mikroskopisi
- 60x (yağ) apokromatik amaç veya benzer bir konfigürasyona sahip başka bir mikroskop kullanarak CCD kamera ile donatılmış ters floresan mikroskop kullanarak Parlak alan (BF) ve floresan (IF) filtreleri altındaki lekeli hücreleri görüntüleyin.
6. RE lokalize proteinleri ve melanozomlar arasındaki örtüklerin nicelemesi:
NOT: Mander'in Fiji yazılımını kullanan proteinler arasındaki örtük katsayısının ölçülmesi için aşağıdaki adımlar izlenir (bağlantıdan serbestçe indirilebilir: https://imagej.net/software/fiji/). TIFF görüntüsünü birden fazla kanalla kullanın.
- Ham görüntüyü açın. Görüntü seçeneğine gidin, Renk | Kanalları bölün ve analiz için iki kanalı kullanın.
- JaCoP eklentisini Eklenti seçeneğinde açın.
- Her iki kanal için de eşiği, tüm parlak noktaların seçilip arka planın ortadan kaldırılması için ayarlayın.
- Analiz seçeneğine gidin, Mander'in çakışma katsayısını almak için M1 ve M2 katsayılarını seçin.
- JACoP Eklentisi'ndeki Analiz Et seçeneğine basın ve Mander'ın çakışma katsayısını görüntüleyen sonucu görün.
7. Geri dönüşüm endozomlarının borulu sayısının ve uzunluğunun nicelleştirilmesi:
NOT: Fiji yazılımını kullanan tübüllerin sayısının ve uzunluğunun ölçülmesi için aşağıdaki adımlar izlenir.
- Ham görüntüyü açma Görüntüye Git seçeneğini belirleyin, Renk | Kanalları bölün ve analiz için istediğiniz kanalı kullanın.
- Görüntü'ye yeniden gidin, | Yaz'ı seçin 8 bit görüntüye dönüştürün.
- Ardından Eklentiler'e gidin, | Çözümle'yi seçin Tüplük. Sigma değerini 0,1075 olarak ayarlayın. Tamam'a basın.
- Görüntü'ye yeniden gidin, | Yaz'ı seçin 8 bit görüntüye dönüştürün.
- Resim | git | ayarlama Eşik (Tüm görüntüler için aynı eşik değerlerini kullanın. Görüntüler yaklaşık olarak eşit yoğunluğa sahip olmalıdır).
- İşlem | git İkili | Maskeye dönüştürün.
- İşlem'e gidin, İkili'yi seçin ve sonra İskeletleştir'i seçin.
- Analiz Et'e gidin, İskelet'i seçin ve İskeleti Analiz Et'i seçin. Sonuç ve çıktı | seçin (a) En büyük kısa yolu hesaplayın | (b) ayrıntılı bilgi göster | (c) etiketli iskeleti görüntüleyin. Tamam'a basın.
NOT: Sonuç tablolanmış biçimde açılır. Ortalama dal uzunluğu sütunu, seçilen hücredeki tüm farklı tübüllerin uzunluğunu gösterir (mikrometre cinsinden değer elde için ölçek ayarlayın). - Hücredeki tübül sayısını elde etmek için Çözümle'ye gidin ve Parçacığı analiz et seçeneğini belirleyin. Tamam'a basın.
NOT: Elde edilen sonuçlarda, sayım sütunu belirli bir hücredeki tübül sayısını gösterir. - Verileri kaydedin ve analiz edin.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
STX13Δ129 mutant lokalizasyonunun melanozomlara ölçülmesi
Fare vahşi tip melanositlerinde STX13 immünofluoresans mikroskopisi, GFP-STX13WT'nin veziküler ve tübüler yapılar olarak lokalize olduğunu ve GFP-STX13Δ129'un hücre yüzeyine ek olarak halka benzeri yapılar olarak lokalize olduğunu göstermiştir (Şekil 1A). Ayrıca hücre içi halka benzeri GFP-STX13Δ129 melanozom proteini TYRP1 (Şekil 1A) ve parlak alan görüntülenmiş melanozomlar (veriler gösterilmez)12 ile kolokalizasyon göstermiştir. Daha önce de gösterildiği gibi, melanozomlarda aşırı ifade edilmiş GFP-STX13WT grubu gözlenir12. GFP-STX13WT ve GFP-STX13Δ129'un melanosomes'a göreli yerelleştirilmesini ölçmek için Fiji'yi kullandık ve JACoP eklentisi ile analiz ettik. TYRP1 ile GFP-STX13Δ129 arasındaki ölçülen Mander'in örtüşme katsayısı (MOC), TYRP1 ile GFP-STX13WT'ye kıyasla yaklaşık 1,5 kat daha yüksektir (Şekil 1B). İlginçtir ki, TYRP1 GFP-STX13Δ129 ile GFP-STX13WT'ye kıyasla 2,9 kat daha yüksek MOC değerleri gösterdi (Şekil 1B). Bu veriler, GFP-STX13Δ129'un melanozomlara lokalizasyonunun, sabit bir durumda GFP-STX13WT'ye kıyasla nispeten daha yüksek olduğunu göstermektedir.
STX13WT yerelleştirmesinin geri dönüşüm endozomlarına ölçülmesi
GFP-STX13WT immünofluoresans mikroskopisi bilinen geri dönüşüm endosomal proteini Rab11 (mCherry-Rab11 olarak ifade edilir) ile kolokalizasyon göstermiştir (Şekil 2A,B). mCherry-Rab11 ile GFP-STX13WT arasındaki ölçülen MOC, GFP-STX13WT ile mCherry-Rab11'e kıyasla yaklaşık 1,4 kat daha yüksektir (Şekil 2B). GFP-STX13WT pozitif endosomal tübüllerin sayısını ve uzunluğunu ölçmek için protokol bölümünde açıklandığı gibi Fiji yazılımını kullandık. mCherry-Rab11 deneylerde pozitif kontrol olarak kullanılmaktadır (Şekil 2). GFP-STX13WT ile transkomositler, mCherry-Rab11'i ifade eden hücrelere kıyasla hücre başına daha yüksek sayıda tübül gösterdi (Şekil 2C üst grafiği, A çubuğunu B çubuğuyla karşılaştırın). Bununla birlikte, tübül sayıları hücrelerde GFP-STX13WT ve mCherry-Rab11'in birlikte ifade edilirken azaltılır (Şekil 2C üst grafiği, A çubuğunu C ile karşılaştırın ve B çubuğunu D ile karşılaştırın). İlginçtir ki, hem GFP-STX13WT hem de mCherry-Rab11 için ortalama tübül uzunluğu (μm), ayrı ayrı veya birlikte ifade eden hücrelerde birbiriyle karşılaştırılabilir (Şekil 2C alt grafik). Birlikte, bu veriler GFP-STX13WT'nin Rab11'e benzer şekilde REs'e yerelleştirme yaptığını göstermektedir.
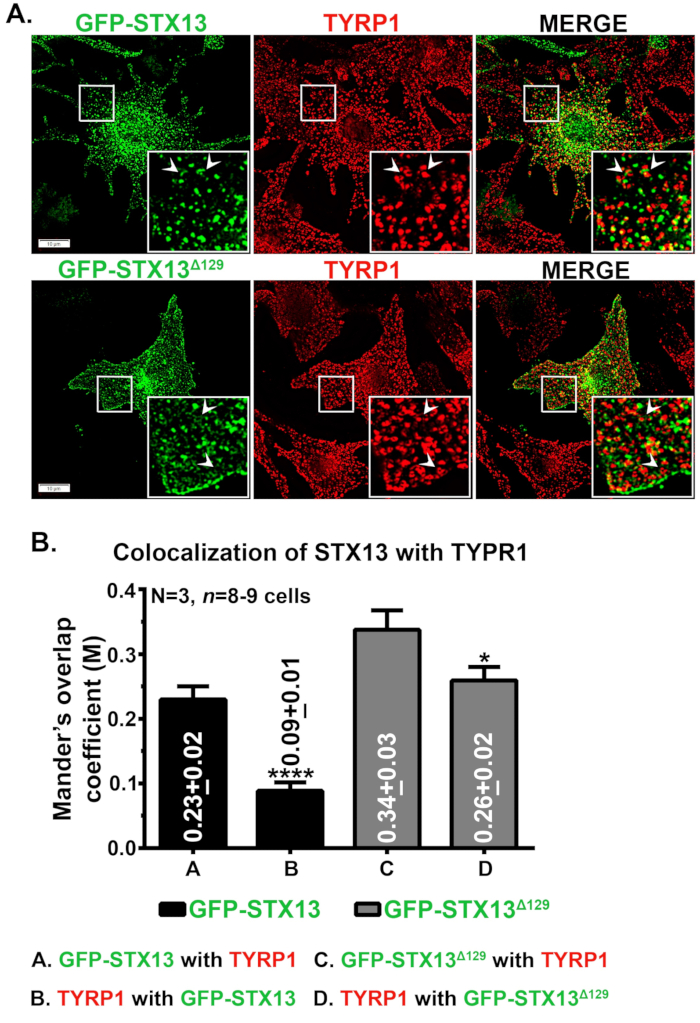
Şekil 1: GFP-STX13WT ve GFP-STX13Δ129'un yabani tip melanositlerdeki melanozomlara lokalizasyonu. (A) Melan-Ink4a-Arf-1 melanositleri GFP-STX13WT ve GFP-STX13Δ129 ile transktalize edildi. Hücreler sabitlendi, anti-TYRP1 antikoru ile boyandı ve daha sonra floresan mikroskopi ile analiz edildi. İç kısımlar beyaz kutulu alanların büyütülmüş görünümüne sahiptir. Oklar, GFP-STX13WTto REs ve GFP-STX13Δ129'un melanozomlara yerelleştirilmesine işaret eder. Ölçek çubukları, 10 μm. (B) STX13 ve TYRP1 arasındaki kolokalizasyonun nicelemesi. Mander'in TYRP1 ile GFP-STX13WT veya GFP-STX13Δ129 arasındaki örtüşme katsayısı (M) ve bunun tersi de arsada ayrı ayrı temsil edilir (ortalama ± S.E.M.). N=3. *p ≤0.05 ve ****p ≤0.0001. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.

Şekil 2: GFP-STX13WT'nin yabani tip melanositlerdeki endozomların geri dönüşümüne lokalizasyonu. (A) Melan-Ink4a-Arf-1 melanositleri GFP-STX13WT ve mCherry-Rab11 ile transkredaklı hale getirildi. Hücreler floresan mikroskopi ile sabitlendi ve analiz edildi. İç kısımlar beyaz kutulu alanların büyütülmüş görünümüne sahiptir. Oklar, GFP-STX13WT'nin mCherry-Rab11 pozitif bölmelere yerelleştirilmesine işaret eder. Ölçek çubukları, 10 μm. (B) STX13 ve Rab11 arasındaki kolokalizasyonun nicelemesi. Mander'in mCherry-Rab11 ile GFP-STX13WT arasındaki örtüştürük katsayısı (M) ve bunun tersi arsada ayrı ayrı temsil edilir (ortalama ± S.E.M.). N=3. s ≤0.001. C. STX13 veya Rab11 pozitif RE'lerin sayı ve uzunluğunun (μm olarak) ölçülmesi. GFP-STX13WT ve mCherry-Rab11'in ortalama tübül/hücre ve ortalama tübül uzunluğu (ortalama ± S.E.M.) arsada ayrı ayrı temsil edilir. N=3. Hücrelerin GFP-STX13WT ve mCherry-Rab11 ile (C) ve (D) olarak transkine olduğunu unutmayın. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Geri dönüşüm endozomları endosit organellerden oluşan bir kohort haline gelerek tüm hücre tiplerinde kargonun hücre yüzeyine geri dönüşümüne aracılık ederler21,22,23,24,25. Melanositler gibi özel hücre tiplerinde, bu organeller kısmen biyogenezleri için kaçakçılık rotalarını melanozomlara yönlendirmektedir3,16,26. Ayrıca, RE'ler erken /sıralama endozomlarında kargo sonrası sıralama oluşturulur ve morfolojik olarak tübüler-veziküler yapılar olarak görünür. Dinamikleri (sayı ve uzunluk) motor proteinler21,22,23,24,25,27,28 dahil olmak üzere çeşitli hücresel faktörlere bağlıdır. Çok az çalışma, Rab11'i melanositler14,19'da REs'i etiketlemek için bir işaretleyici olarak kullandı. Melanosit olmayanlarda, STX13 ve KIF13A, REs13,17,19'u görselleştirmek ve karakterize etmek için Rab11'e ek olarak kullanılmıştır. Bu çalışmada STX13'ü melanositlerdeki RE'leri etiketlemek ve ardından Rab11-pozitif tübüllerle karşılaştırmak için bir işaretleyici olarak kullandık (Şekil 2). İlginçtir ki, STX13'teki (GFP-STX13Δ129) N-terminal silme, SNARE'nin melanozomlara yanlış yönlendirilmesiyle sonuçlanır (Şekil 1). Bu nedenle, GFP-STX13WT ve GFP-STX13Δ129'un melanositlerde lokalizasyonu sırasıyla REs ve melanozomları görselleştirir. Bu proteinler, sabit durum lokalizasyon çalışmaları için ilgili organel belirteçleri olarak kullanılabilir.
Çalışmalar, TYRP1, TYR (tyrosinaz), VAMP7, OCA2, Rab32/38 gibi çeşitli proteinlerin diğer hücre içi organellere ek olarak melanozomlara lokalize olduğunu göstermiştir3,29. Çalışmalarımız, N-terminali silinen STX13 mutantın (GFP-STX13Δ129) melanozomlara ve plazma zarlarına lokalize olduğunu göstermiştir. GFP-STX13Δ129'un, YP'lerden hücre yüzeyine ve LRO'ya yapılan kaçakçılığı incelemek için bir muhabir olarak kullanılabileceğini vardır. Buna karşılık, GFP-STX13WT, Rab11'e benzer şekilde REs'e yerelleştirir. Çalışmalarımız, GFP-STX13WT'nin melanositlerdeki REs'leri işaretlemek için de kullanılabileceğini göstermektedir. GFP-STX13WT'nin Rab11'den daha iyi bir RE işaretleyici olabileceğini öngörüyoruz, çünkü Rab11 aşırı ifade endosomal dinamikleri değiştiriyor. GFP-STX13WT, dinamiklerini sabit durumda incelemek için potansiyel bir RE işaretçisi görevi görür.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Yazarlar çıkar çatışması olmadığını beyan ederler.
Acknowledgments
Bu çalışma Biyoteknoloji Bölümü tarafından desteklendi (BT/PR32489/BRB/10/1786/2019 to SRGS); Bilim ve Mühendislik Araştırma Kurulu (CRG/2019/000281 to SRGS); DBT-NBACD (BT/HRD-NBA-NWB/38/2019-20 to SRGS) ve IISc-DBT ortaklık programı (SRGS'ye). Bölümdeki altyapı DST-FIST, DBT ve UGC tarafından desteklendi. AMB, DBT-JRF (DBT/2015/IISc/NJ-02) tarafından desteklendi.
Materials
| Name | Company | Catalog Number | Comments |
| anti-TYRP1 antibody (TA99) | ATCC | HB-8704 | |
| Fluoromount-G | Southern Biotech | 0100-01 | |
| Lipofectamine 2000 | ThermoFisher Scientific | 11668-500 | |
| Matrigel matrix | BD Biosciences | 356231 | |
| OPTI-MEM | ThermoFisher Scientific | 022600-050 | |
| Phorbol 12-myristate 13-acetate | Sigma-Aldrich | P8139 | |
| RPMI Medium 1640 | ThermoFisher Scientific | 31800-022 |
References
- Dell'Angelica, E. C. The building BLOC(k)s of lysosomes and related organelles. Current Opinion in Cell Biology. 16 (4), 458-464 (2004).
- Raposo, G., Marks, M. S. Melanosomes--dark organelles enlighten endosomal membrane transport. Nature Reviews in Molecular Cell Biology. 8 (10), 786-797 (2007).
- Ohbayashi, N., Fukuda, M. Recent advances in understanding the molecular basis of melanogenesis in melanocytes. F1000Research. 9, F1000 Faculty Rev-608 (2020).
- Theos, A. C., et al. Functions of adaptor protein (AP)-3 and AP-1 in tyrosinase sorting from endosomes to melanosomes. Molecular Biology of the Cell. 16 (11), 5356-5372 (2005).
- Di Pietro, S. M., et al. BLOC-1 interacts with BLOC-2 and the AP-3 complex to facilitate protein trafficking on endosomes. Molecular Biology of the Cell. 17 (9), 4027-4038 (2006).
- Setty, S. R., et al. BLOC-1 is required for cargo-specific sorting from vacuolar early endosomes toward lysosome-related organelles. Molecular Biology of the Cell. 18 (3), 768-780 (2007).
- Delevoye, C., et al. AP-1 and KIF13A coordinate endosomal sorting and positioning during melanosome biogenesis. Journal of Cell Biology. 187 (2), 247-264 (2009).
- Bultema, J. J., Ambrosio, A. L., Burek, C. L., Di Pietro, S. M. BLOC-2, AP-3, and AP-1 proteins function in concert with Rab38 and Rab32 proteins to mediate protein trafficking to lysosome-related organelles. Journal of Biological Chemistry. 287 (23), 19550-19563 (2012).
- Sitaram, A., et al. Differential recognition of a dileucine-based sorting signal by AP-1 and AP-3 reveals a requirement for both BLOC-1 and AP-3 in delivery of OCA2 to melanosomes. Molecular Biology of the Cell. 23 (16), 3178-3192 (2012).
- Nag, S., et al. Rab4A organizes endosomal domains for sorting cargo to lysosome-related organelles. Journal of Cell Science. 131 (18), (2018).
- Dennis, M. K., et al. BLOC-2 targets recycling endosomal tubules to melanosomes for cargo delivery. Journal of Cell Biology. 209 (4), 563-577 (2015).
- Jani, R. A., Purushothaman, L. K., Rani, S., Bergam, P., Setty, S. R. STX13 regulates cargo delivery from recycling endosomes during melanosome biogenesis. Journal Cell Science. 128 (17), 3263-3276 (2015).
- Shakya, S., et al. Rab22A recruits BLOC-1 and BLOC-2 to promote the biogenesis of recycling endosomes. EMBO Reports. 19 (12), 45918 (2018).
- Bowman, S. L., et al. A BLOC-1-AP-3 super-complex sorts a cis-SNARE complex into endosome-derived tubular transport carriers. Journal of Cell Biology. 220 (7), 202005173 (2021).
- Wei, M. L. Hermansky-Pudlak syndrome: a disease of protein trafficking and organelle function. Pigment Cell Research. 19 (1), 19-42 (2006).
- Bowman, S. L., Bi-Karchin, J., Le, L., Marks, M. S. The road to lysosome-related organelles: Insights from Hermansky-Pudlak syndrome and other rare diseases. Traffic. 20 (6), 404-435 (2019).
- Prekeris, R., Klumperman, J., Chen, Y. A., Scheller, R. H. Syntaxin 13 mediates cycling of plasma membrane proteins via tubulovesicular recycling endosomes. Journal of Cell Biology. 143 (4), 957-971 (1998).
- Mahanty, S., et al. Rab9A is required for delivery of cargo from recycling endosomes to melanosomes. Pigment Cell Melanoma Research. 29 (1), 43-59 (2016).
- Delevoye, C., et al. Recycling endosome tubule morphogenesis from sorting endosomes requires the kinesin motor KIF13A. Cell Reports. 6 (3), 445-454 (2014).
- Ha, L., et al. ARF functions as a melanoma tumor suppressor by inducing p53-independent senescence. Proceedings of the National Academy of Science U. S. A. 104 (26), 10968-10973 (2007).
- Soldati, T., Schliwa, M. Powering membrane traffic in endocytosis and recycling. Nature Reviews Molecular Cell Biology. 7 (12), 897-908 (2006).
- Grant, B. D., Donaldson, J. G. Pathways and mechanisms of endocytic recycling. Nature Reviews Molecular Cell Biology. 10 (9), 597-608 (2009).
- Hsu, V. W., Prekeris, R.
- Taguchi, T.
- Goldenring, J. R.
- Delevoye, C., Marks, M. S., Raposo, G. Lysosome-related organelles as functional adaptations of the endolysosomal system. Current Opinion in Cell Biology. 59, 147-158 (2019).
- Hsu, V. W., Bai, M., Li, J. Getting active: protein sorting in endocytic recycling. Nature Reviews in Molecular Cell Biology. 13 (5), 323-328 (2012).
- Desfougeres, Y., D'Agostino, M., Mayer, A. A modular tethering complex for endosomal recycling. Nature Cell Biology. 17 (5), 540-541 (2015).
- Le, L., Sires-Campos, J., Raposo, G., Delevoye, C., Marks, M. S. Melanosome biogenesis in the pigmentation of mammalian skin. Integrated Computational Biology. 61 (4), 1517-1545 (2021).
