

Summary
إعادة تدوير الاندوسومات هي جزء من شبكة أنبوبية endosomal. هنا نقدم طريقة لقياس ديناميات إعادة تدوير الاندوسومات باستخدام GFP-STX13 كعلامة عضوية.
Abstract
إعادة تدوير الاندوسومات (REs) هي العضيات الأنبوبية المركبات المتولدة من الإنسومات المبكرة / الفرز في جميع أنواع الخلايا. تلعب هذه العضيات دورا رئيسيا في التكوين الحيوي للميلانوسومات ، وهي عضوية مرتبطة بالليسوسوم تنتجها الخلايا الصباغية. REs تسليم البضائع الخاصة بالخلايا الصباغية إلى الميلانوسومات المبكرة أثناء تشكيلها. انسداد في جيل REs، لوحظ في العديد من المسوخ من متلازمة هيرمانسكي بودلاك، يؤدي إلى نقص التصبغ من الجلد والشعر والعين. لذلك ، فإن دراسة ديناميكيات (الرجوع إلى عدد وطول) REs مفيدة لفهم وظيفة هذه العضيات في الظروف الطبيعية والمرضية. في هذه الدراسة، ونحن نهدف إلى قياس ديناميات RE باستخدام STX13 SNARE المقيمين.
Introduction
يحدث التمثيل الحيوي لأصباغ الميلانين في الميلانوسومات ، وهي عضوية مرتبطة بالليسوسومات الخاصة بالخلايا الصباغية (LRO) تتعايش مع الليسوسومات التقليدية. يلعب النظام الإندوسيتيكي دورا رئيسيا في التكوين الحيوي للميلانوسومات ، المطلوبة للون البشرة وprotection ضد الإشعاع المؤين1،2،3. خلال هذه العملية، يتم فرز الإنزيمات توليف الميلانين على الاندوسومات في وقت مبكر / الفرز ومن ثم نقلها إلى الميلانوسومات المبكرة من خلال الاندوسومات أنبوبي أو المركبات تسمى إعادة تدوير الانسومات (REs)4،5،6،7،8،9،10. استهداف وانصهار هذه العضيات تنظيم نضوج الميلانوسومات المصطبغة تعمل بكامل طاقتها7،11،12،13،14. العيوب في تشكيل هذه العضيات أو فرز البضائع إلى هذه العضيات تسبب المهق oculocutaneous وغيرها من الأنماط الظاهرية السريرية، لوحظ في متلازمة هيرمانسكي بودلاك15،16.
هنا نصف تقنية بسيطة تعتمد على المجهر لدراسة وتحليل REs. في هذه الطريقة، لقد استفدنا من بروتين ترانسممبران، Qa-SNARE Syntaxin (STX)13 الذي يقيم على إعادة تدوير endosomes17 ودورات بين الاندوسومات الفرز والميلانوسومات في الخلايا الصباغية12،18. علاوة على ذلك ، فإن حذف النطاق التنظيمي غير المنظم N-terminal (أي SynN أو STX13Δ129) يسمح لل SNARE بالتعثر في الميلانوسومات ، والذي يقيس مسار الاتجار إلى الأمام نحو الميلانوسوم12. لقد استخدمنا علامة إعادة تدوير نهايةوسومية معروفة Rab GTPase (Rab)11 في دراساتنا14,19. التصوير الفلوري للبروتينات GFP-STX13WT، GFP-STX13Δ129، mCherry-Rab11، و TYRP1 في الخلايا الصباغية البرية من النوع تليها تحديد كمي لتوطينها النسبي سيوفر طبيعة وديناميات REs بالإضافة إلى استهدافها للميلانوسومات. وهكذا، هذه تقنية بسيطة يمكن استخدامها لتصور وقياس ديناميات REs في الخلايا الصباغية.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
يتضمن البروتوكول بذر الخلايا الصباغية متبوعا بتحول البلازميدات. وتشمل الخطوات الأخرى تثبيت الخلايا، والتتيح المناعي، والتصوير، وتحليلها لقياس طول وعدد ال REs. ويرد أدناه الوصف المفصل للبروتوكول.
1. بذر الخلايا الصباغية الماوس على coverlips المعالجة مسبقا
- معطف coverlips الزجاج في طبق بيتري (أي 4- 5 في طبق 35 ملم) مع مصفوفة غشاء الطابق السفلي المتوسطة (1:20 في المتوسط RPMI كاملة: RPMI + 10٪ الحرارة تعطيل FBS + 1x الجلوتامين + 1x مزيج المضادات الحيوية) وتجفيفه في غطاء محرك السيارة ثقافة الأنسجة لمدة 15 دقيقة. غسل الأغطية مرة واحدة مع برنامج تلفزيوني 1x قبل الاستخدام.
- الحفاظ على الخلايا الصباغية الماوس نوع البرية (الميلان-Ink4a-Arf-1 من C57BL/6J، a/a، Ink4a-Arf-/- mice، وصف in20 ومتاحة في الثقة ترحيب وظيفية الجينوم بنك الخلية) في طبق بيتري (أي، 35 أو 60 مم طبق) تكملها كاملة RPMI المتوسطة.
- غسل الخلايا مرتين مع برنامج تلفزيوني 1x وإضافة 0.5 أو 1 مل من تريبسين-EDTA (0.25٪) حل لخلايا الجربسينج. احتضان الخلايا في 37 درجة مئوية لمدة 2 - 5 دقائق للانفصال عن طبق بيتري.
- إضافة 1 - 2 مل من المتوسط RPMI كاملة، تعليق ومن ثم نقل الخلايا إلى أنبوب الطرد المركزي.
- الطرد المركزي تعليق الخلية في 4 درجة مئوية، 376 × ز لمدة 5 دقائق ومن ثم إعادة إنفاق بيليه في برنامج تلفزيوني 1x.
- كرر خطوة الطرد المركزي وإعادة تثبيت الخلايا في 1 مل من المتوسط RPMI كاملة.
- بذور الخلايا على غشاء الطابق السفلي يغطي المتوسطة المغلفة في التقاء 50-60٪ (ما يقرب من 6 × 105 خلايا على طبق ثقافة الخلية 35 ملم تحتوي على 4-5 coverslips). دائما إضافة 200 nM من PMA (إضافة 5 ميكرولتر من المخزون العامل 40 μM phorbol 12-myristate 13-خلات) إلى تعليق الخلية مطلي في المتوسط RPMI كاملة.
- بعد البذر، واحتضان لوحة في 37 درجة مئوية لمدة 12-24 ساعة.
2. إصابة الخلايا مع البلازميدات STX13
- استخدم الكواشف التالية: pEGFP-C1-STX13WT وpEGFP-C1-STX13Δ129 (موضح في12). mCherry-Rab11، كان هدية طيبة من غراسا رابوسو، معهد كوري، باريس (الموصوفة في المرجع19). مضاد TYRP1 من ATCC (TA99).
- بعد 12 - 24 ساعة من البذر ، ينقل الخلايا مع البلازميدات باستخدام كاشف العدوى القائم على الدهون. للحصول على طبق 35 مم، خذ 5 ميكرولتر من كاشف العدوى في 250 ميكرولتر من المتوسط OPTI-MEM في أنبوب طرد مركزي صغير واخذ ما يقرب من 200 نانوغرام من كل بلازميد في 250 ميكرولتر من OPTI-MEM.
- احتضان الأنابيب التي تحتوي على الحمض النووي وكواشف العدوى لمدة 5 دقائق. مزيج دون ماصة المتكررة (إجمالي حجم سيكون حوالي 500 ميكرولتر). حضانة لمدة 30 دقيقة في درجة حرارة الغرفة. قم بالتنصت اليدوي على الأنبوب كل 10 دقائق لمدة 30 دقيقة في RT.
- أثناء الحضانة، اغسل الخلايا مرتين باستخدام برنامج تلفزيوني 1x، مرة واحدة باستخدام OPTI-MEM ثم أضف 1 مل من OPTI-MEM إلى الخلايا.
- بعد 30 دقيقة من الحضانة ، أضف مزيج الكاشف الحمض النووي transfection إلى الخلايا بطريقة دروبوايز من خلال تغطية الطبق.
- احتضان الخلايا في 37 درجة مئوية لمدة 6 ساعة. يستنشق المتوسطة OPTI-MEM مع كاشف transfection وإضافة المتوسطة RPMI كاملة تكملها مع 200 nM من PMA.
- احتضان الخلايا في 37 درجة مئوية لمدة 48 ساعة.
3. تثبيت الخلايا
ملاحظة: يتم تنفيذ الإجراء التالي خارج غطاء محرك السيارة زراعة الأنسجة.
- بعد 48 ساعة من العدوى، وغسل الخلايا مرتين مع برنامج تلفزيوني 1x ومن ثم إصلاح الخلايا مع 3٪ الفورمالديهايد (أعدت حديثا في برنامج تلفزيوني 1x) لمدة 30 دقيقة.
- بعد التثبيت، اغسل الخلايا مرتين مع برنامج تلفزيوني 1x وتخزين الأغطية في برنامج تلفزيوني 1x حتى مزيد من الاستخدام. بدلا من ذلك، يمكن تركيب الخلايا على الشرائح الزجاجية (انظر أدناه) أو تخزينها عند 4 درجات مئوية.
4. الخلايا المناعية
- إعداد غرفة رطبة: وضع قطعة قطع فيلم البارافين على ورقة تصفية رطبة في طبق بيتري، مغطاة رقائق الألومنيوم.
- إعداد 25 ميكرولتر من محلول الأجسام المضادة الأولية (0.2٪ سابونين في برنامج تلفزيوني 1x، 0.1٪ BSA في برنامج تلفزيوني 1x و 0.02٪ أزيد الصوديوم في برنامج تلفزيوني 1x). إضافة الأجسام المضادة في تخفيف 1:200. إضافة هذا الحل كقطر على فيلم البارافين في غرفة رطبة.
- رفع بعناية coverlip مع ملقط، عكس ذلك على هذا قطرة من محلول تلطيخ الأجسام المضادة الأولية ومن ثم تغطية غطاء الغرفة الرطبة. احتضان في درجة حرارة الغرفة لمدة 30 دقيقة.
- وبالمثل، إعداد حل الأجسام المضادة الثانوية في تخفيف 1:500 ووضعه على فيلم البارافين بجوار coverslip في غرفة رطبة. لتلطيخ النواة، أضف DAPI (1:20,000 إلى 1:30,000) إلى الحل.
- باستخدام ملقط، والتقاط بعناية coverslip من الحل الأساسي للأجسام المضادة وتراجع ثلاث مرات في برنامج تلفزيوني 1x (في كوب زجاجي).
- اضغط على غطاء على ورق الأنسجة لإزالة برنامج تلفزيوني الزائدة على coverlip. وضعه على حل تلطيخ الأجسام المضادة الثانوية في غرفة رطبة ولا يعرضها للضوء بسبب وجود الأجسام المضادة الموسومة الفلورسنت في الحل.
- احتضان غطاء مرة أخرى لمدة 30 دقيقة في درجة حرارة الغرفة. الرجاء دوما الاحتفاظ ملاحظة جانب coverslip يحتوي على الخلايا خلال هذه الخطوات.
- بعد الحضانة ، التقط بعناية الغطاء من محلول الأجسام المضادة الثانوي ثم تراجعه ثلاث مرات في برنامج تلفزيوني 1x. علاوة على ذلك، اضغط على غطاء الغطاء على ورق الأنسجة لإزالة برنامج تلفزيوني الزائد على غطاء.
- ضع 12 ميكرولتر من كاشف التصاعد Fluoromount-G على شريحة زجاجية وضع بعناية غطاء ملون (مواجهة نحو الزجاج) على كاشف التركيب. عكس الشريحة الزجاجية على ورق الأنسجة ثم اضغط بلطف.
5. مضان المجهر من الخلايا
- صورة الخلايا الملطخة تحت برايت فيلد (BF) والفلورسينس (IF) مرشحات باستخدام مجهر مضان مقلوب مجهز بكاميرا CCD باستخدام هدف 60x (النفط) apochromatic أو أي مجهر آخر مع تكوين مماثل.
6. تحديد التداخل بين البروتينات المحلية RE والميلانوسومات:
ملاحظة: يتم اتباع الخطوات التالية للقياس الكمي لمعامل التداخل بين البروتينات باستخدام برنامج فيجي (يمكن تنزيله بحرية من الرابط: https://imagej.net/software/fiji/). استخدم صورة TIFF مع قنوات متعددة.
- افتح الصورة الخام. انتقل إلى الخيار صورة، حدد اللون | تقسيم القنوات، واستخدام القناتين للتحليل.
- افتح البرنامج المساعد JACoP في خيار المكون الإضافي.
- تعيين عتبة لكلا القناتين بحيث يتم تحديد جميع النقاط المضيئة ويتم القضاء على الخلفية.
- انتقل إلى خيار التحليل، حدد معاملات M1 و M2 للحصول على معامل التداخل في Mander.
- اضغط على الخيار تحليل في البرنامج المساعد JACoP وانظر النتيجة التي تعرض معامل التداخل Mander.
7. تحديد كمي لإعادة تدوير الاندوسومات 'أنبوبي عدد وطول:
ملاحظة: تتبع الخطوات التالية من أجل تحديد عدد وطول الأنابيب باستخدام برنامج فيجي.
- افتح الخيار "الانتقال إلى الصورة" للصورة، وحدد اللون | تقسيم القنوات واستخدام القناة المطلوبة للتحليل.
- انتقل إلى الصورة مرة أخرى، وحدد نوع | تحويل إلى صورة 8 بت.
- ثم انتقل إلى الإضافات، حدد تحليل | أنبوبية. تعيين قيمة سيغما في 0.1075 . اضغط على موافق.
- انتقل إلى الصورة مرة أخرى، وحدد نوع | تحويل إلى صورة 8 بت.
- انتقل إلى | الصور ضبط | العتبة (استخدم نفس قيم العتبة لكافة الصور. يجب أن يكون للصور كثافة متساوية تقريبا).
- انتقل إلى | العملية | ثنائي تحويل إلى قناع.
- انتقل إلى عملية، حدد ثنائي ثم حدد Skeletonize.
- انتقل إلى تحليل، حدد الهيكل العظمي واختر تحليل الهيكل العظمي. في النتيجة والمخرجات | حدد ( أ) حساب أكبر أقصر مسار | (ب) إظهار معلومات مفصلة | (ج) عرض الهيكل العظمي المسمى. اضغط على موافق.
ملاحظة: سيتم فتح النتيجة في تنسيق جدولة. يظهر عمود متوسط طول الفرع طول جميع الأنابيب المختلفة في الخلية المحددة (مقياس مجموعة للحصول على قيم بالميكرومتر). - للحصول على عدد الأنابيب في الخلية، انتقل إلى تحليل، وحدد تحليل الجسيمات الخيار. اضغط على موافق.
ملاحظة: في النتائج التي تم الحصول عليها، يظهر عمود العد عدد الأنابيب في تلك الخلية المعينة. - حفظ البيانات وتحليلها.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
القياس الكمي لتوطين متحول STX13Δ129 إلى الميلانوسومات
أظهر المجهر المناعي من STX13 في الخلايا الصباغية نوع الماوس البرية GFP-STX13WT المترجمة كهياكل المركبات والأنابيب وGFP-STX13Δ129 المترجمة كهياكل تشبه حلقة بالإضافة إلى سطح الخلية (الشكل 1A). علاوة على ذلك، أظهرت GFP-STX13Δ129 الشبيهة بالحلقة داخل الخلايا كولوكاليزم مع بروتين الميلانوسوم TYRP1 (الشكل 1A) والميلانوسومات المصورة ذات المجال الساطع (البيانات غير المعروضة)12. كما هو مبين من قبل، لوحظ مجموعة من GFP-STX13WT مفرط التعبير في الميلانوسوم12. لقياس التعريب النسبي لGFP-STX13WT وGFP-STX13Δ129 إلى الميلانوسومات، استخدمنا فيجي وحللنا مع البرنامج المساعد JACoP. معامل التداخل المقاس بين GFP-STX13Δ129 مع TYRP1 أعلى بنحو 1.5 مرة مقارنة ب GFP-STX13WT مع TYRP1 (الشكل 1B). ومن المثير للاهتمام أن TYRP1 أظهر قيم MOC أعلى بمقدار 2.9 طيات مع GFP-STX13Δ129 مقارنة ب GFP-STX13WT (الشكل 1B). تشير هذه البيانات إلى أن توطين GFP-STX13Δ129 إلى الميلانوسومات أعلى نسبيا مقارنة ب GFP-STX13WT في حالة ثابتة.
تحديد كمية توطين STX13WT إلى الاندوسومات المعاد تدويرها
أظهر المجهر المناعي من GFP-STX13WT الكولوكال مع البروتين الإنضومالي المعروف إعادة التدوير Rab11 (أعرب عن mCherry-Rab11) (الشكل 2A، B). إن MOC المقاسة بين GFP-STX13WT مع mCherry-Rab11 أعلى بنحو 1.4 أضعاف مقارنة ب mCherry-Rab11 مع GFP-STX13WT (الشكل 2B). لقياس عدد وطول الأنابيب endosomal GFP-STX13WT إيجابية، استخدمنا برنامج فيجي كما هو موضح في قسم البروتوكول. يستخدم mCherry-Rab11 كتحكم إيجابي في التجارب (الشكل 2). وأظهرت الخلايا الصباغية التي تم تحويلها مع GFP-STX13WT عددا أكبر من الأنابيب لكل خلية مقارنة بالخلايا التي تعبر عن mCherry-Rab11 (الرسم البياني العلوي الشكل 2C، قارن الشريط A مع الشريط B). ومع ذلك، يتم تقليل أرقام أنبوبي على التعبير المشترك من GFP-STX13WT و mCherry-Rab11 في الخلايا (الشكل 2C الرسم البياني العلوي، قارن شريط A مع C، وشريط B مع D). ومن المثير للاهتمام أن متوسط طول أنبوبي (μm) لكل من GFP-STX13WT و mCherry-Rab11 مماثل لبعضها البعض في الخلايا التي تعبر بشكل فردي أو معا (الرسم البياني السفلي الشكل 2C). معا، تشير هذه البيانات إلى أن GFP-STX13WT مترجمة إلى REs على أنها مشابهة ل Rab11.
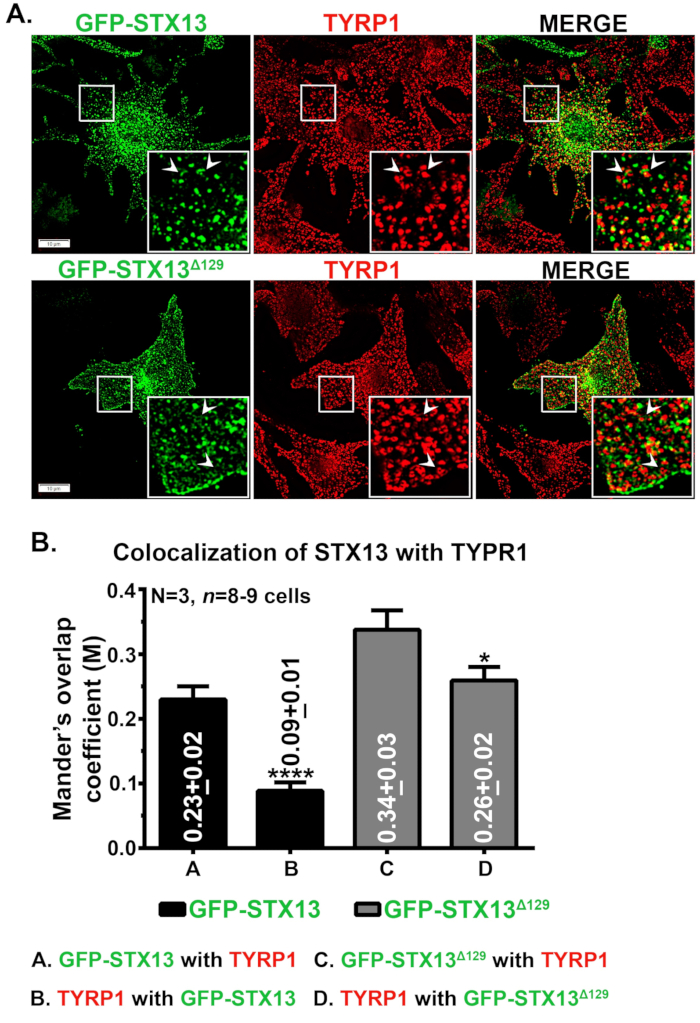
الشكل 1: توطين GFP-STX13WT وGFP-STX13Δ129 إلى الميلانوسومات في الخلايا الصباغية البرية من النوع. (أ) تم تحويل الخلايا الصباغية Melan-Ink4a-Arf-1 مع GFP-STX13WT وGFP-STX13Δ129. تم إصلاح الخلايا، ملطخة بالأجسام المضادة TYRP1 ومن ثم تحليلها عن طريق المجهر الفلوري. يتم تكبير عرض Insets للمناطق المربعة البيضاء. تشير الأسهم إلى توطين GFP-STX13WTto REs و GFP-STX13Δ129 إلى الميلانوسومات. أشرطة المقياس، 10 ميكرومتر. (ب) تحديد حجم الكولوسيوم بين STX13 و TYRP1. معامل التداخل بين ماندر (M) بين GFP-STX13WT أو GFP-STX13Δ129 مع TYRP1 والعكس صحيح يتم تمثيله (متوسط ± S.E.M.) بشكل منفصل في المؤامرة. N = 3. *p ≤0.05 و ****p ≤0.0001. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.

الشكل 2: تم تحويل توطين GFP-STX13WT لإعادة تدوير الاندوسومات في الخلايا الصباغية البرية. (أ) الخلايا الصباغية Melan-Ink4a-Arf-1 مع GFP-STX13WT و mCherry-Rab11. تم إصلاح الخلايا وتحليلها عن طريق المجهر الفلوري. يتم تكبير عرض Insets للمناطق المربعة البيضاء. تشير الأسهم إلى توطين GFP-STX13WT إلى مقصورات mCherry-Rab11 الإيجابية. أشرطة المقياس، 10 ميكرومتر. (ب) تحديد حجم الكولوسيوم بين STX13 و Rab11. يتم تمثيل معامل التداخل (M) بين GFP-STX13WT مع mCherry-Rab11 والعكس بالعكس (متوسط ± S.E.M.) بشكل منفصل في المؤامرة. N = 3. ص ≤0.001. ج- تحديد عدد وطول (في ميكرومتر) من STX13- أو REs إيجابية Rab11. يتم تمثيل متوسط عدد الأنابيب / الخلية ومتوسط طول أنبوبي من GFP-STX13WT و mCherry-Rab11 (متوسط ± S.E.M.) بشكل منفصل في المؤامرة. N = 3. لاحظ أن يتم نقل الخلايا مع GFP-STX13WT و mCherry-Rab11 في (C) و (D). يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
إعادة تدوير الاندوسومات هي مجموعة من العضيات الانسطية، وأنها تتوسط في إعادة تدوير البضائع إلى سطح الخلية في جميع أنواع الخلايا21،22،23،24،25. في أنواع الخلايا المتخصصة مثل الخلايا الصباغية، تحول هذه العضيات جزئيا طريق الاتجار بها نحو الميلانوسومات لنشأتها الحيوية3،16،26. وعلاوة على ذلك، يتم إنشاء REs بعد فرز البضائع على الوسومات المبكرة / الفرز وتظهر بشكل مورفولوجي كهياكل أنبوبية المركبات. تعتمد ديناميكياتها (العدد والطول) على العديد من العوامل الخلوية، بما في ذلك البروتينات الحركية21,22,23,24,25,27,28. استخدمت دراسات قليلة Rab11 كعلامة لتسمية REs في الخلايا الصباغية14،19. في حين أنه في الخلايا غير الصباغية، تم استخدام STX13 و KIF13A بالإضافة إلى Rab11 لتصور وتوصيف REs13،17،19. في هذه الدراسة، استخدمنا STX13 كعلامة لتسمية REs في الخلايا الصباغية تليها مقارنة مع الأنابيب الإيجابية Rab11 (الشكل 2). ومن المثير للاهتمام، N-حذف المحطة الطرفية في STX13 (GFP-STX13Δ129) النتائج في سوء تقدير من الفخ إلى الميلانوسومات (الشكل 1). وهكذا، فإن توطين GFP-STX13WT و GFP-STX13Δ129 في الخلايا الصباغية تصور REs والميلانوسومات، على التوالي. يمكن استخدام هذه البروتينات كعلامات العضيات المعنية لدراسات توطين الحالة الثابتة.
وقد أظهرت الدراسات أن العديد من البروتينات مثل TYRP1، TYR (التيروزيناسي)، VAMP7، OCA2، Rab32/38 توطين الميلانوسومات بالإضافة إلى العضيات داخل الخلايا الأخرى3،29. أظهرت دراساتنا أن N-المحطة الطرفية حذف STX13 متحولة (GFP-STX13Δ129) يحط من الميلانوسومات وغشاء البلازما. ونحن نفترض أن GFP-STX13Δ129 يمكن أن تستخدم ربما كمراسل لدراسة الاتجار من REs إلى سطح الخلية وLRO. في المقابل، ترجمة GFP-STX13WT إلى REs مشابهة ل Rab11. أوضحت دراساتنا أنه يمكن أيضا استخدام GFP-STX13WT لوضع علامات على REs في الخلايا الصباغية. ونحن نتوقع أن GFP-STX13WT قد يكون أفضل علامة RE من Rab11 منذ التعبير المفرط Rab11 يغير ديناميات endosomal. بالإجمال، يعمل GFP-STX13WT كعلامة RE محتملة لدراسة ديناميكياتها في حالة ثابتة.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
ويعلن صاحبا البلاغ أنه ليس بينهما تضارب في المصالح.
Acknowledgments
وقد دعم هذا العمل قسم التكنولوجيا الحيوية (BT/PR32489/BRB/10/1786/2019 إلى SRGS)؛ مجلس بحوث العلوم والهندسة (CRG/2019/000281 إلى SRGS)؛ DBT-NBACD (BT/HRD-NBA-NWB/38/2019-20 إلى SRGS) وبرنامج شراكة IISc-DBT (إلى SRGS). تم دعم البنية التحتية في القسم من قبل DST-FIST و DBT و UGC. تم اعتماد AMB من قبل DBT-JRF (DBT/2015/IISc/NJ-02).
Materials
| Name | Company | Catalog Number | Comments |
| anti-TYRP1 antibody (TA99) | ATCC | HB-8704 | |
| Fluoromount-G | Southern Biotech | 0100-01 | |
| Lipofectamine 2000 | ThermoFisher Scientific | 11668-500 | |
| Matrigel matrix | BD Biosciences | 356231 | |
| OPTI-MEM | ThermoFisher Scientific | 022600-050 | |
| Phorbol 12-myristate 13-acetate | Sigma-Aldrich | P8139 | |
| RPMI Medium 1640 | ThermoFisher Scientific | 31800-022 |
References
- Dell'Angelica, E. C. The building BLOC(k)s of lysosomes and related organelles. Current Opinion in Cell Biology. 16 (4), 458-464 (2004).
- Raposo, G., Marks, M. S. Melanosomes--dark organelles enlighten endosomal membrane transport. Nature Reviews in Molecular Cell Biology. 8 (10), 786-797 (2007).
- Ohbayashi, N., Fukuda, M. Recent advances in understanding the molecular basis of melanogenesis in melanocytes. F1000Research. 9, F1000 Faculty Rev-608 (2020).
- Theos, A. C., et al. Functions of adaptor protein (AP)-3 and AP-1 in tyrosinase sorting from endosomes to melanosomes. Molecular Biology of the Cell. 16 (11), 5356-5372 (2005).
- Di Pietro, S. M., et al. BLOC-1 interacts with BLOC-2 and the AP-3 complex to facilitate protein trafficking on endosomes. Molecular Biology of the Cell. 17 (9), 4027-4038 (2006).
- Setty, S. R., et al. BLOC-1 is required for cargo-specific sorting from vacuolar early endosomes toward lysosome-related organelles. Molecular Biology of the Cell. 18 (3), 768-780 (2007).
- Delevoye, C., et al. AP-1 and KIF13A coordinate endosomal sorting and positioning during melanosome biogenesis. Journal of Cell Biology. 187 (2), 247-264 (2009).
- Bultema, J. J., Ambrosio, A. L., Burek, C. L., Di Pietro, S. M. BLOC-2, AP-3, and AP-1 proteins function in concert with Rab38 and Rab32 proteins to mediate protein trafficking to lysosome-related organelles. Journal of Biological Chemistry. 287 (23), 19550-19563 (2012).
- Sitaram, A., et al. Differential recognition of a dileucine-based sorting signal by AP-1 and AP-3 reveals a requirement for both BLOC-1 and AP-3 in delivery of OCA2 to melanosomes. Molecular Biology of the Cell. 23 (16), 3178-3192 (2012).
- Nag, S., et al. Rab4A organizes endosomal domains for sorting cargo to lysosome-related organelles. Journal of Cell Science. 131 (18), (2018).
- Dennis, M. K., et al. BLOC-2 targets recycling endosomal tubules to melanosomes for cargo delivery. Journal of Cell Biology. 209 (4), 563-577 (2015).
- Jani, R. A., Purushothaman, L. K., Rani, S., Bergam, P., Setty, S. R. STX13 regulates cargo delivery from recycling endosomes during melanosome biogenesis. Journal Cell Science. 128 (17), 3263-3276 (2015).
- Shakya, S., et al. Rab22A recruits BLOC-1 and BLOC-2 to promote the biogenesis of recycling endosomes. EMBO Reports. 19 (12), 45918 (2018).
- Bowman, S. L., et al. A BLOC-1-AP-3 super-complex sorts a cis-SNARE complex into endosome-derived tubular transport carriers. Journal of Cell Biology. 220 (7), 202005173 (2021).
- Wei, M. L. Hermansky-Pudlak syndrome: a disease of protein trafficking and organelle function. Pigment Cell Research. 19 (1), 19-42 (2006).
- Bowman, S. L., Bi-Karchin, J., Le, L., Marks, M. S. The road to lysosome-related organelles: Insights from Hermansky-Pudlak syndrome and other rare diseases. Traffic. 20 (6), 404-435 (2019).
- Prekeris, R., Klumperman, J., Chen, Y. A., Scheller, R. H. Syntaxin 13 mediates cycling of plasma membrane proteins via tubulovesicular recycling endosomes. Journal of Cell Biology. 143 (4), 957-971 (1998).
- Mahanty, S., et al. Rab9A is required for delivery of cargo from recycling endosomes to melanosomes. Pigment Cell Melanoma Research. 29 (1), 43-59 (2016).
- Delevoye, C., et al. Recycling endosome tubule morphogenesis from sorting endosomes requires the kinesin motor KIF13A. Cell Reports. 6 (3), 445-454 (2014).
- Ha, L., et al. ARF functions as a melanoma tumor suppressor by inducing p53-independent senescence. Proceedings of the National Academy of Science U. S. A. 104 (26), 10968-10973 (2007).
- Soldati, T., Schliwa, M. Powering membrane traffic in endocytosis and recycling. Nature Reviews Molecular Cell Biology. 7 (12), 897-908 (2006).
- Grant, B. D., Donaldson, J. G. Pathways and mechanisms of endocytic recycling. Nature Reviews Molecular Cell Biology. 10 (9), 597-608 (2009).
- Hsu, V. W., Prekeris, R.
- Taguchi, T.
- Goldenring, J. R.
- Delevoye, C., Marks, M. S., Raposo, G. Lysosome-related organelles as functional adaptations of the endolysosomal system. Current Opinion in Cell Biology. 59, 147-158 (2019).
- Hsu, V. W., Bai, M., Li, J. Getting active: protein sorting in endocytic recycling. Nature Reviews in Molecular Cell Biology. 13 (5), 323-328 (2012).
- Desfougeres, Y., D'Agostino, M., Mayer, A. A modular tethering complex for endosomal recycling. Nature Cell Biology. 17 (5), 540-541 (2015).
- Le, L., Sires-Campos, J., Raposo, G., Delevoye, C., Marks, M. S. Melanosome biogenesis in the pigmentation of mammalian skin. Integrated Computational Biology. 61 (4), 1517-1545 (2021).
