

Summary
子宮内膜のリサイクルは、内膜管状ネットワークの一部です。ここでは、GFP-STX13をオルガネラマーカーとして使用して、エンドーソームのリサイクルのダイナミクスを定量化する方法を紹介します。
Abstract
循環性の子宮体(R)は、すべての細胞タイプの早期/選別用の子宮間から生成される管状小胞性小器官である。これらのオルガネラは、メラノサイトによって産生されるリソソーム関連小器官であるメラノソームの生物形成において重要な役割を果たす。Rは、メラノサイト特有の貨物を形成中に早期メラノソームに提供します。レスキー・プドラック症候群のいくつかの変異体で観察される、レージの生成における閉塞は、皮膚、髪、および眼の色素沈着をもたらす。したがって、レジェのダイナミクス(数と長さを参照)を研究することは、正常および疾患状態におけるこれらのオルガネラの機能を理解するのに有用である。本研究では、居住SNARE STX13を用いてREダイナミクスを測定することを目指す。
Introduction
メラニン色素の生合成は、メラノソーム、従来のリソソームと共存するメラノサイト特異的リソソーガネラ(LRO)で起こる。エンドサイト系は、色素体の生物形成において重要な役割を果たし、放射線電離放射線に対する皮膚色と光保護に必要な1,2,3です。この過程で、メラニン合成酵素は早期/選別エンドーソームで選別され、その後、循環性エンドーソーム(RE)4,5,6,7,8,9,10と呼ばれる管状または小胞性子宮体を介して早期メラノソームに輸送されます。これらのオルガネラの標的化および融合は、完全に機能性色素性メラノソーム7,11,12,13,14の成熟を調節する。これらのオルガネラまたはこれらのオルガネラへの貨物選別の形成における欠陥は、オキュロチュネアアルビニズムおよび他の臨床表現型を引き起こし、ヘルマンスキー・プドラク症候群15,16で観察された。
ここでは、Rを研究し、分析するための単純な顕微鏡ベースの手法について説明します。この方法では、endoソーム17のリサイクルに存在する膜貫通タンパク質であるQa-SNAREシンコシンシン(STX)13と、メラノサイトにおけるエンドーソームとメラノソーム間のサイクルを利用しています12,18。また、N末端非構造化制御ドメイン(すなわちSynNまたはSTX13Δ129)を削除すると、SNAREはメラノソームに立ち往生し、メラノソーム12に向かう前方の入稿経路を測定する。私たちの研究では、既知の循環型内経マーカーRab GTPase(Rab)11を使用しています14,19。タンパク質GFP-STX13WT、GFP-STX13Δ129、mCherry-Rab11、およびIRP1のタンパク質の蛍光イメージングは、その後、それらの相対的な局在化の定量化に続いて、メラノソームへの標的化に加えて、REの性質とダイナミクスを提供する。このように、これはメラノサイトのRのダイナミクスを可視化し、測定するために使用できる簡単な技術です。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
プロトコルは、プラスミドのトランスフェクションに続いてメラノサイトの播種を伴う。さらにステップには、細胞の固定、免疫染色、イメージング、および分析が含まれ、レの長さと数を測定する。プロトコルの詳細な説明は以下の通りです。
1. 前処理されたカバーリップ上のマウスメラノサイトの播種
- ガラスカバーリップをペトリ皿(すなわち、35mm皿の4-5)に基膜マトリックス培地(完全RPMI培地では1:20:RPMI + 10%熱不活性化FBS + 1xグルタミン+1x抗生物質ミックス)でコーティングし、ティッシュ培養フードで15分間乾燥させます。使用前に1倍PBSでカバーリップを1回洗ってください。
- 野生型マウスメラノサイト(C57BL/6Jのメランインク4a-Arf-1、 a/a、 Ink4a-Arf--マウス 、20 年に記載され、ウェルカムトラスト機能ゲノミクスセルバンクで入手可能)をペトリ皿(すなわち、35または60mm皿)で維持する。
- 1x PBSで細胞を2回洗浄し、トリプシン細胞をトリプシンするためのトリプシンEDTA(0.25%)溶液を0.5または1mL添加します。37°Cで2〜5分間細胞をインキュベートし、ペトリ皿から剥離します。
- 完全なRPMI培地の1〜2mLを加え、中断し、遠心分離管に細胞を移します。
- 細胞懸濁液を4°Cで遠心し、376 x g で5分間、ペレットを1x PBSで再懸濁させた。
- 遠心分離工程を繰り返し、完全RPMI培地の1mLで細胞を再懸濁します。
- 50~60%の合流度で基膜培地コーティングカバーリップの上に細胞を播種します(4~5個のカバーリップを含む35mm細胞培養皿の約6 x 105 細胞)。完全なRPMI媒体のメッキされた細胞懸濁液にPMAの200 nM(作業ストック40 μMの40 μMの40 μLの12-myristate 13-アセテートを加える)を加えます。
- 播種後、37°Cでプレートを12〜24時間インキュベートする。
2. STX13プラスミドを用いて細胞をトランスフェクションする
- 以下の試薬を使用してください: pEGFP-C1-STX13WT および pEGFP-C1-STX13Δ129 (説明 12) 。mCherry-Rab11は、グラサ・ラポソ、キュリー研究所、パリからの親切な贈り物でした(参考19に記載されています)。ATCCからの抗TYRP1抗体(TA99)。
- 12〜24時間の播種後、脂質系トランスフェクション試薬を用いてプラスミドで細胞をトランスフェクトする。35mmの皿の場合、マイクロ遠心分離管内のOPTI-MEM培地の250μLにトランスフェクション試薬を5μLとし、OPTI-MEMの250μLで各プラスミドの約200ngを取ります。
- DNAとトランスフェクション試薬を含むチューブを5分間インキュベートします。繰り返しピペットを使わずに混合します(総体積は約500μLになります)。室温で30分間インキュベートします。RTで30分間、10分ごとにチューブの手タップを行います。
- インキュベーション中に、細胞を1x PBSで2回、OPTI-MEMで1回洗浄し、細胞に1mLのOPTI-MEMを加えます。
- インキュベーションのポスト30分、皿を覆うことによって細胞にトランスフェクション試薬-DNAミックスを加える。
- 37°Cで6時間培養する。トランスフェクション試薬でOPTI-MEM培地を吸引し、PMAの200 nMを添加した完全なRPMI培地を添加する。
- 37°Cで48時間培養します。
3. 細胞の固定
注:以下の手順は、組織培養フードの外側で行われます。
- 48時間のトランスフェクションの後、1x PBSで細胞を2回洗浄し、3%ホルムアルデヒド(1x PBSで作りたての)で細胞を30分間固定します。
- 固定後、細胞を1x PBSで2回洗浄し、さらに使用するまでカバーリップを1倍PBSに保管します。あるいは、セルはガラススライド(下記参照)に取り付けたり、4°Cで保存することができます。
4. 細胞の免疫染色
- 湿気の多いチャンバーを準備する:ペトリ皿の湿った濾紙にパラフィンフィルムカットピースを置き、アルミニウム箔で覆います。
- 一次抗体溶液25μL(1x PBSでは0.2%サポニン、1x PBSでは0.1%BSA、1x PBSでは0.02%のアジドナトリウム)を調製します。1:200の希釈時に抗体を加える。湿気の多いチャンバーのパラフィンフィルムに滴としてこの溶液を追加します。
- 慎重に鉗子でカバースリップを持ち上げ、一次抗体染色溶液のこの滴でそれを反転させ、湿潤チャンバーの蓋を覆う。室温で30分間インキュベートします。
- 同様に、2次抗体溶液を1:500の希釈で調製し、湿気の多いチャンバーのカバースリップの隣のパラフィンフィルムに置きます。核を染色するには、DAPI (1:20,000 ~ 1:30,000) を溶液に加えます。
- 鉗子を使用して、慎重に一次抗体溶液からカバースリップを拾い、それを1x PBS(ガラスビーカー)に3回浸します。
- ティッシュペーパーのカバースリップをタップして、カバースリップの余分なPBSを取り除きます。湿気の多いチャンバー内の二次抗体染色液の上に置き、溶液中に蛍光タグ付き抗体が存在するため、光にさらしません。
- カバースリップを室温で30分間再びインキュベートします。これらの手順を通して、セルを持つカバースリップの側面を常にメモしてください。
- インキュベーション後、二次抗体溶液からカバースリップを慎重に取り出し、1x PBSに3回浸します。さらに、ティッシュペーパーのカバースリップをタップして、カバースリップ上の余分なPBSを取り除きます。
- フッ素マウントG実装試薬12μLをガラススライドに配置し、取り付け試薬の上にステンドカバースリップ(ガラスに向かって向いている)を慎重に配置します。ティッシュペーパーのガラススライドを反転し、軽く押します。
5. 細胞の蛍光顕微鏡
- 60x(油)のアポクロマティック目的または同様の構成を有する他の顕微鏡を用いたCCDカメラを搭載した反転蛍光顕微鏡を使用して、明視野(BF)および蛍光(IF)フィルター下の染色細胞を画像化します。
RE局所タンパク質とメラノソーム間の重複の定量化:
注: フィジーソフトウェアを使用したタンパク質間のマンダーの重複係数の定量化については、以下の手順に従います(リンクから自由にダウンロード可能:https://imagej.net/software/fiji/)。TIFF イメージを複数のチャネルで使用します。
- 生の画像を開きます。[イメージ] オプションに移動し、[色] |チャネルを分割し、分析に 2 つのチャネルを使用します。
- 「プラグイン」オプションでJACoPプラグインを開きます。
- すべての明るいスポットが選択され、背景が削除されるように、両方のチャネルのしきい値を設定します。
- [解析]オプションに移動し、マンダーの重なり係数を取得するには、M1とM2係数を選択します。
- JACoPプラグインの 分析オプション を押して、マンダーの重なり係数を表示する結果を確認します。
7. エンドソームの管数と長さのリサイクルの定量化:
注: フィジーソフトウェアを使用した管の数と長さの定量化については、次の手順に従います。
- 未加工の画像を開く 画像に移動 ]オプションを選択し、[ カラー]|チャネルを分割 し、目的のチャネルを使用して解析します。
- もう一度 [イメージ] に移動し、[ 種類] | 8 ビット イメージに変換します。
- 次にプラグインに移動 し、[分析] を選択 |チューブネス。 シグマ 値を 0.1075 に設定します。 [OK] を押します。
- もう一度 [イメージ] に移動し、[ 種類] を選択| 8 ビットイメージに変換します。
- イメージ |に移動|を調整するしきい値(すべての画像に同じしきい値を使用します。画像の強度はほぼ等しい必要があります)。
- プロセス |に移動バイナリ|マスクに変換します。
- [プロセス] に移動し、[バイナリ] を選択して、[スケルトン化] を選択します。
- [解析]に移動し、[スケルトン]を選択して、[スケルトンを解析]を選択します。結果と出力|選択(a)最大最短経路|を計算(b)は、(c)ラベル付きスケルトンを表示|詳細情報を表示します。[OK] を押します。
注: 結果は、表形式で開きます。[平均分岐長]の列には、選択されたセル内のすべての異なる細管の長さが表示されます(マイクロメートルで値を取得するためのスケールを設定します)。 - セル内のチューブの数を取得するには、[ 解析] に移動し、[ パーティクルの分析 ] オプションを選択します。 [OK] を押します。
注: 取得した結果では、count 列に特定のセル内の管の数が表示されます。 - データを保存して分析します。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
STX13Δ129変異体のメラノソームへの定量化
マウス野生型メラノサイトにおけるSTX13の免疫蛍光顕微鏡検査は、GFP-STX13WTを細胞表面に加えて環状構造として局在した小胞および管状構造として局在し、GFP-STX133WTを示した(図1A)。また、細胞内リング様GFP-STX13Δ129は、メラノソームタンパク質のTYRP1(図1A)と明視野画像メラノソーム(データは示さない)と共局在化を示した。前に示したように、過剰発現GFP-STX13WTのコホートはメラノソーム12で観察される。メラノソームへのGFP-STX13WTとGFP-STX13Δ129の相対的な局在を測定するために、我々はフィジーを使用し、JACoPプラグインで分析しました。TYRP1 を有する GFP-STX13Δ129 間の測定マンダーのオーバーラップ係数 (MOC) は、TYRP1 を搭載した GFP-STX13WT と比較して約 1.5 倍高くなっています (図 1B)。興味深いことに、TYRP1はGFP-STX133WTと比較してGFP-STX13Δ129で2.9倍高いMOC値を示した(図1B)。これらのデータは、メラノソームに対するGFP-STX13Δ129の局在化が、安定した状態でのGFP-STX13WTと比較して比較的高いことを示している。
循環型のendosomesへのSTX13WT局在化の定量化
GFP-STX13WTの免疫蛍光顕微鏡は、既知の循環型内毒素Rab11(mCherry-Rab11として表される)と共局在化を示した(図2A、B)。mCherry-Rab11 を搭載した GFP-STX13WT 間の測定された MOC は、GFP-STX13WT を搭載した mCherry-Rab11 と比較して約 1.4 倍高くなっています (図 2B)。GFP-STX13WT陽性の内皮細管の数と長さを測定するために、プロトコルセクションで説明されているようにフィジーのソフトウェアを使用しました。mCherry-Rab11は実験において陽性対照として用いられる(図2)。GFP-STX13WTにトランスフェクトされたメラノサイトは、mCherry-Rab11を発現する細胞と比較して細胞当たりの細管数が多いことを示した(図2Cトップグラフ、バーAとバーBを比較)。ただし、細胞内のGFP-STX13WTとmCherry-Rab11の共発現時に細管数が減少します(図2C上グラフ、バーAとC、バーBとDを比較)。興味深いことに、GFP-STX13WTとmCherry-Rab11の両方の平均細管長(μm)は、個別にまたは一緒に発現する細胞において互いに同等である(図2C下のグラフ)。これらのデータを組み合わせることで、GFP-STX13WTはRab11と同様にREにローカライズすることを示唆しています。
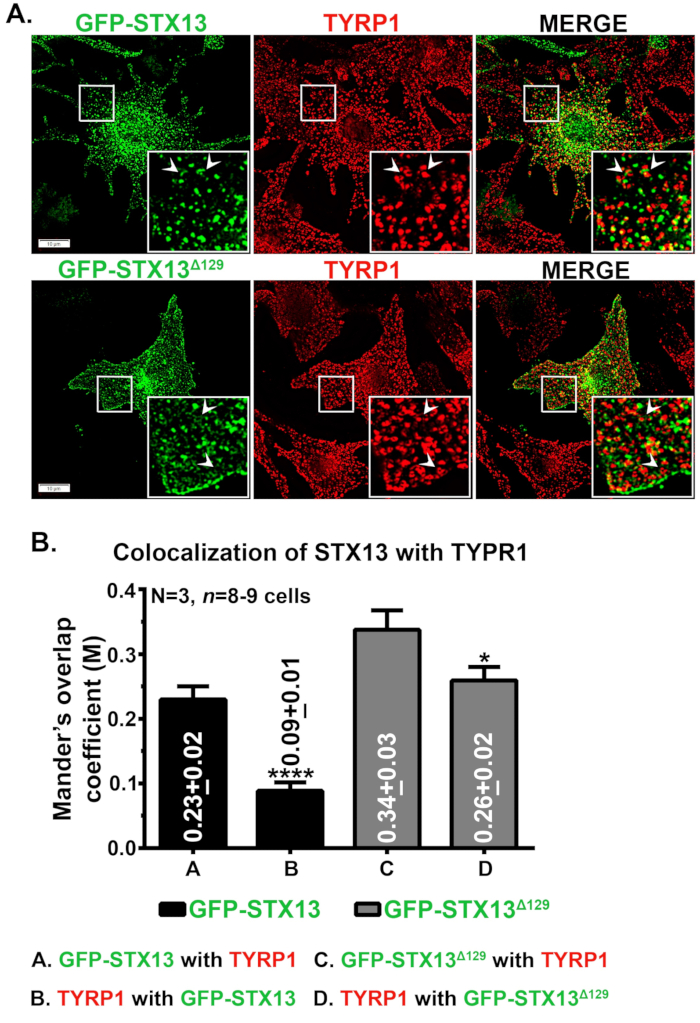
図1:野生型メラノサイトにおけるメラノソームに対するGFP-STX13WTおよびGFP-STX13Δ129の局在化(A)メラノサイトをGFP-STX13WTおよびGFP-STX13Δ129でトランスフェクトした。細胞を固定し、抗TYRP1抗体で染色し、次いで蛍光顕微鏡で分析した。インセットは、白いボックス化された領域の拡大表示です。矢印は、メラノソームへのGFP-STX13WTTO REsおよびGFP-STX13Δ129の局在化を指す。スケールバー、10 μm。(B) STX13 と TYRP1 間の共局在化の定量化GFP-STX13WT または TYRP1 と GFP-STX13Δ129 の間のマンダーのオーバーラップ係数 (M) と、その逆の場合は、プロットで別々に表されます (S.E.M.) を ±別々に表します。N=3。*p ≤0.05 および ****p ≤0.0001.この図の大きなバージョンを表示するには、ここをクリックしてください。

図2:野生型メラノサイト中のエンドーソームをリサイクルするためのGFP-STX13WTの局在化. (A)メラノサイトをGFP-STX13WTおよびmCherry-Rab11でトランスフェクトした。細胞を固定し、蛍光顕微鏡で分析した。インセットは、白いボックス化された領域の拡大表示です。矢印は、mCherry-Rab11-プラスコンパートメントへのGFP-STX13WTの局在化を指しています。スケールバー、10 μm。(B) STX13とRab11の共局化の定量化MCherry-Rab11 との GFP-STX13WT との間のマンダーのオーバーラップ係数 (M) とその逆は、プロットで別々に表されます (平均 ± S.E.M.)。N=3。p ≤0.001.STX13またはRab11陽性REの数と長さ(μm)の定量化。GFP-STX13WTおよびmCherry-Rab11の尿管/細胞および平均細管長の平均数は、プロット内で別々に表される(平均±S.E.M.)N=3。細胞はGFP-STX13WTおよびmCherry-Rab11(C)および(D)でトランスフェクトされることに注意してください。この図の大きなバージョンを表示するには、ここをクリックしてください。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
エンドソームのリサイクルは、エンドサイトオルガネラのコホートであり、全ての細胞型で細胞表面への貨物のリサイクルを仲介します21,22,23,24,25。メラノサイトなどの特殊な細胞型では、これらのオルガネラは、その生物形成のためのメラノソームに向かって彼らの人身売買ルートを部分的に転用する3,16,26。さらに、早期/仕分け用の遠分化の貨物分類後に生成されたレは、形態学的に管状-小胞構造として現れる。それらのダイナミクス(数および長さ)は、運動タンパク質21、22、23、24、25、27、28を含むいくつかの細胞因子に依存している。メラノサイト14,19でRを標識するマーカーとしてRab11を使用した研究はほとんどありませんでした。一方、非メラノサイトでは、STX13およびKIF13Aは、REs13,17,19を視覚化し、特徴付けるためにRab11に加えて使用されています。本研究では、STX13をマーカーとしてメラノサイトでRを標識し、続いてRab11陽性の尿管と比較しました(図2)。興味深いことに、STX13(GFP-STX13Δ129)におけるN末端欠失は、メラノソームへのSNAREの誤局在化をもたらす(図1)。このように、メラノサイトにおけるGFP-STX13WTおよびGFP-STX13Δ129の局在化は、それぞれ、REsとメラノソームを可視化する。これらのタンパク質は、定常局在化研究のためにそれぞれのオルガネラマーカーとして使用することができる。
研究は、TYRP1、TYR(チロシナーゼ)、VAMP7、OCA2、Rab32/38などのいくつかのタンパク質が、他の細胞内小器官に加えてメラノソームに局地化することを示している3,29。我々の研究は、N末端がSTX13変異体(GFP-STX13Δ129)がメラノソームおよび形質膜に局地化することを示した。我々は、GFP-STX13Δ129が、おそらく、細胞表面およびLROへのEからの人身売買を研究するためのレポーターとして使用することができると仮定する。これに対し、GFP-STX13WT は Rab11 と同様に、R にローカライズします。我々の研究は、GFP-STX13WTがメラノサイトのレをマーキングするためにも使用できることを示した。Rab11過剰発現は内皮動態を変化させるため、GFP-STX13WTはRab11よりも優れたREマーカーである可能性があると予測しています。全体として、GFP-STX13WTは、定常状態でのダイナミクスを研究するための潜在的なREマーカーとして機能します。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
著者らは、利益相反はないと宣言している。
Acknowledgments
この作業は、バイオテクノロジー省(BT/PR32489/BRB/10/1786/2019からSRGS)によって支えられた。理工学研究委員会(CRG/2019/000281 SRGS)DBT-NBACD(BT/HRD-NBA-NWB/38/2019-20からSRGS)とIISc-DBTパートナーシッププログラム(SRGSへ)。この部門のインフラストラクチャは、DST-FIST、DBT、および UGC によってサポートされていました。AMB は DBT-JRF (DBT/2015/IISc/NJ-02) によってサポートされました。
Materials
| Name | Company | Catalog Number | Comments |
| anti-TYRP1 antibody (TA99) | ATCC | HB-8704 | |
| Fluoromount-G | Southern Biotech | 0100-01 | |
| Lipofectamine 2000 | ThermoFisher Scientific | 11668-500 | |
| Matrigel matrix | BD Biosciences | 356231 | |
| OPTI-MEM | ThermoFisher Scientific | 022600-050 | |
| Phorbol 12-myristate 13-acetate | Sigma-Aldrich | P8139 | |
| RPMI Medium 1640 | ThermoFisher Scientific | 31800-022 |
References
- Dell'Angelica, E. C. The building BLOC(k)s of lysosomes and related organelles. Current Opinion in Cell Biology. 16 (4), 458-464 (2004).
- Raposo, G., Marks, M. S. Melanosomes--dark organelles enlighten endosomal membrane transport. Nature Reviews in Molecular Cell Biology. 8 (10), 786-797 (2007).
- Ohbayashi, N., Fukuda, M. Recent advances in understanding the molecular basis of melanogenesis in melanocytes. F1000Research. 9, F1000 Faculty Rev-608 (2020).
- Theos, A. C., et al. Functions of adaptor protein (AP)-3 and AP-1 in tyrosinase sorting from endosomes to melanosomes. Molecular Biology of the Cell. 16 (11), 5356-5372 (2005).
- Di Pietro, S. M., et al. BLOC-1 interacts with BLOC-2 and the AP-3 complex to facilitate protein trafficking on endosomes. Molecular Biology of the Cell. 17 (9), 4027-4038 (2006).
- Setty, S. R., et al. BLOC-1 is required for cargo-specific sorting from vacuolar early endosomes toward lysosome-related organelles. Molecular Biology of the Cell. 18 (3), 768-780 (2007).
- Delevoye, C., et al. AP-1 and KIF13A coordinate endosomal sorting and positioning during melanosome biogenesis. Journal of Cell Biology. 187 (2), 247-264 (2009).
- Bultema, J. J., Ambrosio, A. L., Burek, C. L., Di Pietro, S. M. BLOC-2, AP-3, and AP-1 proteins function in concert with Rab38 and Rab32 proteins to mediate protein trafficking to lysosome-related organelles. Journal of Biological Chemistry. 287 (23), 19550-19563 (2012).
- Sitaram, A., et al. Differential recognition of a dileucine-based sorting signal by AP-1 and AP-3 reveals a requirement for both BLOC-1 and AP-3 in delivery of OCA2 to melanosomes. Molecular Biology of the Cell. 23 (16), 3178-3192 (2012).
- Nag, S., et al. Rab4A organizes endosomal domains for sorting cargo to lysosome-related organelles. Journal of Cell Science. 131 (18), (2018).
- Dennis, M. K., et al. BLOC-2 targets recycling endosomal tubules to melanosomes for cargo delivery. Journal of Cell Biology. 209 (4), 563-577 (2015).
- Jani, R. A., Purushothaman, L. K., Rani, S., Bergam, P., Setty, S. R. STX13 regulates cargo delivery from recycling endosomes during melanosome biogenesis. Journal Cell Science. 128 (17), 3263-3276 (2015).
- Shakya, S., et al. Rab22A recruits BLOC-1 and BLOC-2 to promote the biogenesis of recycling endosomes. EMBO Reports. 19 (12), 45918 (2018).
- Bowman, S. L., et al. A BLOC-1-AP-3 super-complex sorts a cis-SNARE complex into endosome-derived tubular transport carriers. Journal of Cell Biology. 220 (7), 202005173 (2021).
- Wei, M. L. Hermansky-Pudlak syndrome: a disease of protein trafficking and organelle function. Pigment Cell Research. 19 (1), 19-42 (2006).
- Bowman, S. L., Bi-Karchin, J., Le, L., Marks, M. S. The road to lysosome-related organelles: Insights from Hermansky-Pudlak syndrome and other rare diseases. Traffic. 20 (6), 404-435 (2019).
- Prekeris, R., Klumperman, J., Chen, Y. A., Scheller, R. H. Syntaxin 13 mediates cycling of plasma membrane proteins via tubulovesicular recycling endosomes. Journal of Cell Biology. 143 (4), 957-971 (1998).
- Mahanty, S., et al. Rab9A is required for delivery of cargo from recycling endosomes to melanosomes. Pigment Cell Melanoma Research. 29 (1), 43-59 (2016).
- Delevoye, C., et al. Recycling endosome tubule morphogenesis from sorting endosomes requires the kinesin motor KIF13A. Cell Reports. 6 (3), 445-454 (2014).
- Ha, L., et al. ARF functions as a melanoma tumor suppressor by inducing p53-independent senescence. Proceedings of the National Academy of Science U. S. A. 104 (26), 10968-10973 (2007).
- Soldati, T., Schliwa, M. Powering membrane traffic in endocytosis and recycling. Nature Reviews Molecular Cell Biology. 7 (12), 897-908 (2006).
- Grant, B. D., Donaldson, J. G. Pathways and mechanisms of endocytic recycling. Nature Reviews Molecular Cell Biology. 10 (9), 597-608 (2009).
- Hsu, V. W., Prekeris, R.
- Taguchi, T.
- Goldenring, J. R.
- Delevoye, C., Marks, M. S., Raposo, G. Lysosome-related organelles as functional adaptations of the endolysosomal system. Current Opinion in Cell Biology. 59, 147-158 (2019).
- Hsu, V. W., Bai, M., Li, J. Getting active: protein sorting in endocytic recycling. Nature Reviews in Molecular Cell Biology. 13 (5), 323-328 (2012).
- Desfougeres, Y., D'Agostino, M., Mayer, A. A modular tethering complex for endosomal recycling. Nature Cell Biology. 17 (5), 540-541 (2015).
- Le, L., Sires-Campos, J., Raposo, G., Delevoye, C., Marks, M. S. Melanosome biogenesis in the pigmentation of mammalian skin. Integrated Computational Biology. 61 (4), 1517-1545 (2021).
