The luminometer output file contains raw measurements for both firefly and Renilla luciferase proteins. This raw format is compatible with the “3’LIFE – single plate analysis” and “3’LIFE – multiplate analysis” spreadsheets available from the Mangone lab website (www.mangonelab.com). The single plate analysis spreadsheet automatically calculates firefly/Renilla ratio, normalizes each miRNA to the appropriate negative control, and normalizes repression values across each plate. This spreadsheet automatically identifies wells with low Renilla luciferase signal, highlights wells that exhibit repression compared to the negative control, and provides measures of repression across the entire plate (Figure 2). See 24 for detailed explanation of statistical analysis.
Multiple replicates can be analysed using the “multiplate analysis” spreadsheet. Each replicate is compared side by side, and statistical measures of the data are automatically calculated (Figure 3). In addition to comparing replicates with the “Normalized Repression” columns, the user can compare repression between the two miRNAs under the “miRNA#1/miRNA#2” columns. This measure divides the repression index for each miRNA for each replicate. This measure can indicate erroneous values from the luciferase assay (for example abnormally high or low readings with the negative control, see Figure 3, row A9, Rep #3), and wells where the repression index may not indicate substantial repression, but that do exhibit significant differences between the miRNAs. While this measure may not be used directly to indicate a miRNA target, it is useful for identifying outliers, problematic wells, or patterns in the data that are not solely attributed to direct miRNA regulation.
The Repression Index (RI) is used to call a putative miRNA target, with lower values corresponding to higher relative repression. The threshold for calling putative targets is based on the level of stringency required by the researcher, but combining the RI with 3’UTRs that display statistically significant p-values (p <0.05) will indicate high confidence targets (see Figure 2 Rows B8 and B12).
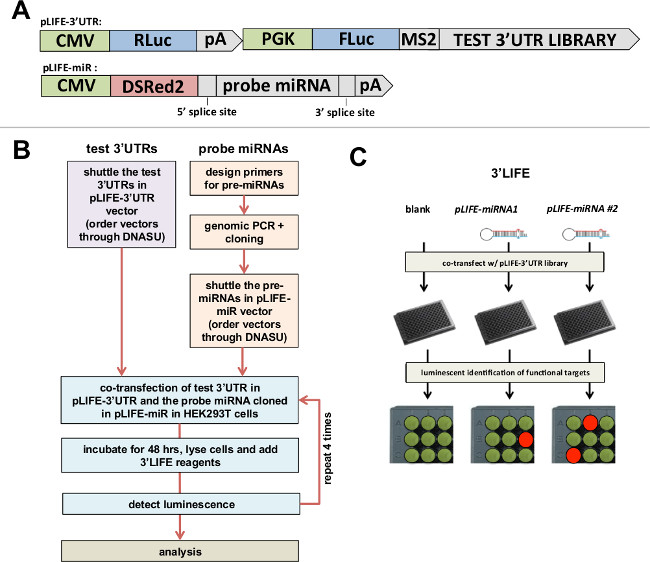
Figure 1: 3’LIFE Assay. (A) Gateway-compatible vectors used in the 3’LIFE assay. Top: The luciferase gene (FLuc) is fused to the test 3’UTR, while the Renilla luciferase gene (RLuc) is fused to the unspecific SV40 pA 3’UTR as control. Bottom: The RFP- miRNA-intron vector – The probe pre-miRNA, plus ~400 nucleotides within its genomic locus (to recapitulate endogenous miRNA processing), is cloned within an intron to allow its co-expression with DSRed2 fluorochrome. Both vectors are publically available (Seiler et al., 2013). (B) Flow chart of the 3’LIFE Assay. (C) 3’LIFE Pipeline: The dual-luciferase vector containing the test 3’UTR with or without the miRNA vectors are co-transfected into HEK293 cells in 96-well plates. The interaction between the miRNA and a bona fide 3’UTR target will lower the relative luminescence in specific wells (exemplified by the orange spot in the experimental plate).
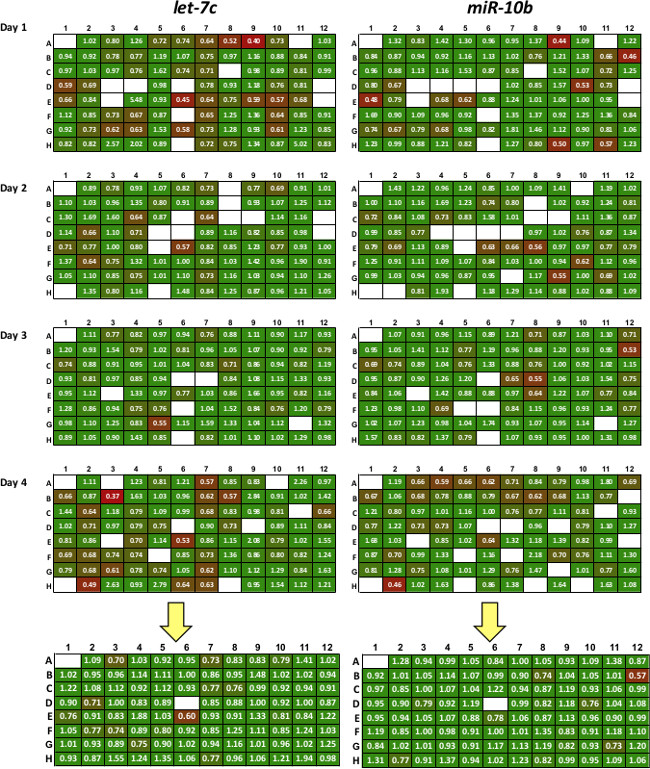
Figure 2: Sample of data produced with 3’LIFE assay. Each probe miRNA is tested in quadruplicate (replicates 1-4). Colors represent repression levels, with red colors indicating strong miRNA/3’UTR interaction. All replicates are averaged to produce high-quality putative targets shown in the summary plate below the yellow arrow. White box represent controls, failed transfections or wells with low transfection efficiency.
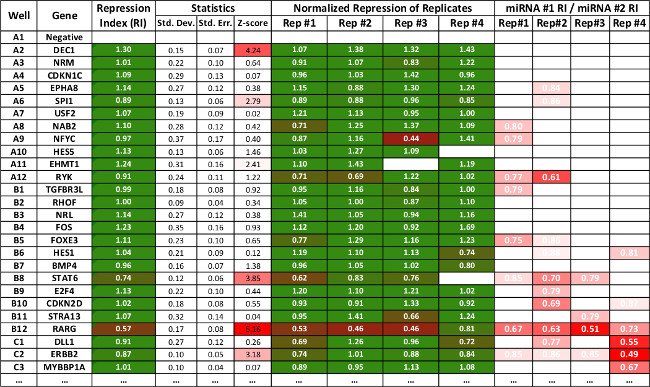
Figure 3: Table representing summary data of a subset of interactions produced using the 3’LIFE spreadsheet. The repression values are as in Figure 2. The software calculates standard deviation, standard error and z-score for each interaction. Statistically significant interactions are marked in red. The last four rows show relative repression of one miRNA to the other, and is used as secondary indicator to compare repression between two different miRNAs. The spreadsheet can be downloaded from www.mangonelab.com
| Firefly luciferase buffer reagents | Final concentration (1x) |
| Glycylglycine | 25 mM |
| KxPO4 (pH 7.8) | 15 mM |
| MgSO4 | 15 mM |
| DTT (store at 4º) | 1 mM |
| EGTA | 4 mM |
| ATP* | 2 mM |
| Beetle luciferin* | 250 μM |
Table 1: Stock firefly luciferase reagents: 10x Stock solutions of Glycylglycine, KxPO4, MgSO4, DTT and EGTA can be prepared separately and stored prior to buffer reconstitution. 100x Beetle Luciferin (firefly luciferase substrate) can be stored by dissolving 50 mg luciferin in 7.134 ml H20 (25 mM). Aliquot 105 μl/plate of dissolved Beetle luciferin into tubes and store at -80 ºC. Per Promega technical support, this should be stable for >6 months, but may be light sensitive. NOTE: EGTA will not go into solution at neutral pH. Slowly add NaOH to EGTA until it dissolves completely.*Reagents added to final buffer immediately prior to the luciferase assay
| Renilla luciferase buffer reagents | Final concentration (1x) |
| NaCl | 1.1 M |
| Na2EDTA | 2.2 mM |
| KH2PO4 | .22 M |
| NaN3 | 1.3 mM |
| BSA* | .44 mg/ml |
| Coelenterazine* | 2.5 μM |
Table 2: Stock Renilla luciferase buffer reagents All the reagents except BSA and Coelenterazine can be mixed at a 1x concentration and stored at room temperature. Coelenterazine can be dissolved in acidified methanol and aliquoted per plate. Acidify methanol by adding HCl to final concentration of 5 mM (<3 pH). Dissolve 250 μg coelenterazine in 2.36 ml acidified methanol (250 μM) aliquot 105 ul/plate. The mix is stable for at least 6 months but may be light sensitive. *Reagents added to buffer immediately before luciferase assay.
